|
||||
|
|
Часть 2. Популяционная экология Популяционная экология – это важнейший элемент общей экологии, так как, по определению Ч. Кребса (Krebs, 1985), «экология – наука о взаимодействиях, определяющих распространение и обилие организмов». Объектами популяционной экологии чаще всего являются виды, имеющие важное хозяйственное значение (вредители сельского и лесного хозяйства, объекты промысла, переносчики опасных заболеваний и т.д.), но иногда это и просто массовые виды или виды редкие, нуждающиеся в охране. Вопросы экологии популяций «стояли за» содержанием предшествующих глав по аутэкологии. Вид –широкая совокупность особей с ареалом, часто простирающимся на десятки тысяч квадратных километров. И по этой причине он адаптируется к условиям среды не целиком, а отдельными популяциями, которые оказываются в разных условиях. Глава 6. Общая характеристика популяции Популящия – совокупность особей одного вида, имеющих общее жизненное пространство и тип взаимоотношений друг с другом. Особи популяции различаются между собой по возрасту и виталитету (т.е. жизненной силе), который может определяться генетически, фенетически, а чаще – сочетанием этих факторов. 6.1. Определение популяции Существует два подхода к пониманию популяции: генетический и экологический. При генетическом подходе под популяцией понимают группу особей одного вида, имеющих общий генофонд, т.е. все особи, которые потенциально могут скрещиваться и обмениваться генами. Такие популяции называются менделевскими. Однако генетическое понимание популяции оказалось неудобным для экологов, так как установить действительно ли особи, обитающие на определенной территории, имеют общий генофонд (т.е. могут скрещиваться) – достаточно трудно, а подчас и просто невозможно. Поэтому в экологии используется иное, прагматическое (т.е. удобное для работы) понятие популяции. Так А.М. Гиляров определяет популяцию следующим образом: «Популяция – любая способная к самовоспроизведению совокупность особей одного вида, более или менее изолированная в пространстве и времени от других аналогичных совокупностей того же вида» (1990, с. 38). При этом сплошь и рядом популяции изолированы «менее, чем более» и переходят друг в друга. Поэтому очень часто их границы экологи устанавливают условно. Кроме неизбежной условности границ популяций, которые устанавливаются с учетом биологического пространства и биологического времени, изучение популяций осложняется разнообразием их внутренней природы, зависящей от биологии организмов. Например по-разному устроены популяции унитарных и модулярных организмов. У первых появлению каждого организма-генета предшествует половой процесс. У вторых из одного генета возможно появление многих десятков и тысяч генетических копий (раметов), которые получаются путем простого деления. К модулярным организмам относятся некоторые животные (мшанки, кораллы, губки, гидроиды, колониальные асцидии) и большинство растений, у которых модулем-метамером является часть побега (стебель с листом и почкой) или целый побег (у клональных растений – злаков, осоковых и некоторых других). Ряд существенных различий, которые нужно учитывать при популяционных исследованиях, имеют популяции растений и животных. Главное отличие заключается в том, что обладающие подвижностью животные могут более активно реагировать на складывающиеся условия внешней среды, избегая неблагоприятных стечений обстоятельств или рассредоточиваясь по территории для компенсации снижения запаса ресурса на единицу площади. Подвижность облегчает им и защиту от хищников. Растения прикреплены («заякорены») к почве и должны реагировать на меняющиеся абиотические (засуха, засоление и т.д.) или биотические (хищники, паразиты, конкуренция с более сильным партнером) условия за счет морфологических и физиолого-биохимических адаптаций. Контрольные вопросы1. Чем отличается определение популяции генетиками от поределения экологов? 2. Перечислите основные отличительные черты популяций растений и животных. 3. Какие организмы относятся к модулярным? 6.2. Конкуренция особей в популяции В силу того, что популяции разнообразны, различаются и взаимодействия особей, входящих в их состав. Поскольку в большинстве случаев популяции обладают способностью к экспоненциальному росту плотности (см. 7.3), суммарная потребность особей, входящих в популяцию, в ресурсах, как правило, больше, чем имеется их в наличии (хотя бы в некоторые периоды жизни особей). По этой причине основным типом взаимодействия особей в популяции является конкуренция, т.е. соревнование за потребление ресурса, которого не хватает. Конкуренция может быть симметричной (конкурирующие особи оказывают одинаковое влияние друг на друга) или асимметричной (влияние особей друг на друга различается по силе). М. Бигон и др. (1989) подчеркивают следующие особенности конкуренции особей в популяции: 1. Конкуренция снижает скорость роста особей, может замедлять их развитие, снижать плодовитость и в итоге – уменьшать вклад в следующие поколения. Количество потомков конкретной особи тем меньше, чем жестче условия конкуренции и чем меньше досталось ей ресурсов. Добавим к этому, что конкуренция может значительно уменьшать размеры растений (особенно эксплерентов). Этот способ реагирования на загущение Дж. Харпер (1973) назвал пластичностью. Размер растений-эксплерентов в зависимости от плотности популяции может меняться в тысячи раз. 2. В большинстве случаев особи конкурируют за ресурсы: каждая особь получает то ограниченное количество ресурсов, которое не было потреблено ее конкурентами. Такая конкуренция называется эксплуатационной. Реже происходит конкуренция за физическое пространство, когда особи «механически» препятствуют друг другу в получении ресурса, скажем, охрана подвижными животными своей территории. Такие отношения называются интерференцией. Поскольку интерференция всегда сопровождается эксплуатацией ресурсов, ее крайне трудно отличить от эксплуатационной конкуренции. В то же время эксплуатационная конкуренция частично не сопровождается интерференцией (т.е. организмы могут потреблять общий ресурс при «мирном» сосуществовании). 3. Разные особи обладают разной конкурентной способностью. Несмотря на то, что все особи популяции потенциально равноценны (идет постоянное выравнивание их генофонда за счет гибридизации), в природе равноценности особей не наблюдается. Популяции гетерогенны как генетически (в их составе могут быть особи с разными признаками – экотипы), так и фенотипически (особи различаются по возрасту и виталитету). При этом животные разных экотипов в силу подвижности могут расходиться по разным местообитаниям, а растения лишены такой возможности. По этой причине генотипическая неоднородность популяций животных, как правило, ниже, чем популяций растений. На исход конкуренции влияет «лотерея», т.е. шанс первым попасть в лучшие или худшие условия среды (микросайты – пятна, различающиеся по благоприятности среды). Особи, которые попали в лучшие условия и (или) начали развиваться раньше, также имеют конкурентные преимущества. Генетические различия, микровариация условий среды и «лотерея» создают предпосылки для дифференциации конкурентной мощности отдельных особей, т.е. разделения их на сильных и слабых, что приводит к асимметричной конкуренции, которая с возрастом особей усиливается (сильный становится еще сильнее, а слабый – слабее, так как ресурсов для него остается все меньше). В итоге асимметричной конкуренции происходит снижение плотности популяции: слабые растения гибнут, а слабые животные мигрируют в местообитания с более низким уровнем конкуренции. Контрольные вопросы1. Приведите примеры симметричной и асимметричной конкуренции. 2. Что такое интерференция? 3. По каким причинам происходит дифференциация конкурентных способностейи особей в популяции? 6.3. Другие формы взаимоотношений особей в популяции Кроме конкуренции возможны и другие формы отношений особей в популяциях – нейтральность (если ресурсов так много и особей так мало, что они практически не мешают друг другу) и положительные взаимоотношения. Взаимовыгодные (или выгодные для части особей) отношения между животными общеизвестны: забота родителей о потомстве, формирование больших семейных групп, стадный образ жизни, коллективная оборона от врагов и т.д. «Караваны» птиц, выстраивающихся в шеренги, клинья, уступы и др., позволяют крыльям отдельных особей в силу аэродинамических эффектов обретать большую подъемную силу (в коллективе летать легче). Есть мнение, что гидродинамические преимущества получают и плывущие стаей рыбы. Однако благоприятные эффекты скопления особей неодинаково выгодны для всего «коллектива». Особенно полезны они для социально сильных животных, которые могут пробиться в центр скопления. У них всегда меньше вероятность быть съеденными хищниками, которые в первую очередь нападают на более слабые особи у периферии скопления. Значительно менее известна роль взаимопомощи у растений. Растения, высеянные группой, развиваются лучше, так как в этом случае у них легче формируется симбиоз с грибами и бактериями микоризы и ризосферы (так называемый«эффект группы»). Именно «эффект группы» был теоретической предпосылкой для разработки «гнездового метода» посева кукурузы и посадки дуба, который пропагандировался сторонниками Т. Лысенко. Однако в силу того, что кратковременное взаимное благоприятствование сменяется острой конкуренцией за ресурсы, при которой происходит взаимное угнетение особей и часть из них гибнет (т.е. напрасно теряются семена или посадочный материал), «гнездовой метод» не прижился на практике. При высеве семян группой со временем формируется так называемый «эффект корыта»: растения в центре группы развиваются хуже, чем расположенные у краев гнезд. Описаны случаи срастания корней у деревьев, при этом часть пластических веществ переходит от более сильного растения к более слабому, возможна передача питательных веществ от одного растения другому через микоризы (см. 8.6). Совместно произрастающие растения эффективнее опыляются насекомыми, так как повышается вероятность переноса пыльцы с цветков одного растения на другое и, кроме того, яркое цветовое пятно из нескольких цветущих и выделяющих ароматические вещества растений лучше привлекает насекомых, чем одно растение. Возможны явления взаимопомощи растений при «коллективной обороне» от фитофагов, проявляющих чрезмерно высокую активность и способных серьезно повредить растениям. В этом случае после начала активного поедания фитофагами в растениях происходят биохимические реакции и повышается концентрация веществ, снижающих их поедаемость (цианидов и др.). Описаны случаи, когда подвергшиеся нападению фитофагов особи выделяли в атмосферу сигнальные вещества (сигнал «меня едят»), которые вызывали повышение образования цианидов у тех особей, которые еще не повреждены. В антропогенных экосистемах уровень конкуренции в популяциях регулирует сам человек, например подбирая нормы высева полевых культур или густоту посадки саженцев в садах и лесопосадках. При этом в большинстве случаев норма высева оказывается несколько завышенной, что позволяет за счет более интенсивной конкуренции снизить потенциальную возможность развития в посевах сорных растений. Подобным образом регулируется конкуренция между карпами при их промышленном разведении в садках. Во многих случаях, дабы не допустить конкуренции, животных содержат в отдельных клетках (песцы, куры, утки и т.д.). Контрольные вопросы1. Приведите примеры взаимопомощи в популяциях животных. 2. В каких случаях проявляется взаимопомощь в популяциях растений? 3. Приведите примеры регулирования плотности популяций человеком. 6.4. Размер популяции и ее структура в пространстве Размер популяции – это количество входящих в нее особей. Он является результирующей взаимодействия биотического потенциала вида и сопротивления среды (рис. 12). 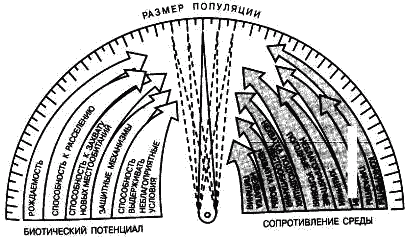 Рис. 12. Факторы, определяющие размер популяции. Сопротивление среды – это комплекс неблагоприятных факторов абиотической и биотической среды, которые воздействуют на организмы. Биотический потенциал – это способность организма преодолевать сопротивление среды. Сравнительно редко удается определить абсолютный размер популяции как общее число особей (т.е. ее численность). Это возможно только для крупных и немногочисленных, обычно находящихся на грани исчезновения видов, обитающих на открытых пространствах саванн, пустынь, травяных болот (львов, слонов, бегемотов и т.д.). В этом случае используется прямой («поголовный») пересчет числа особей, обычно с воздуха. Возможен прямой пересчет особей в небольших популяциях растений (например, венерина башмачка, мамонтова дерева и др.). В остальных случаях размер популяции определяется выборочным методом через плотность популяции – количество особей, приходящихся на единицу площади. Поскольку в разных частях пространства, занимаемого популяцией, ее плотность может различаться, то определяют среднее значение из нескольких учетов. Размер и число учетов-проб, а также достоверность получаемого среднего арифметического определяется в соответствии с требованиями математической статистики. Затем при необходимости можно определить численность популяции путем умножения плотности на занимаемую популяцией площадь. Учет плотности популяций в зависимости от особенностей изучаемого вида проводится разными методами: подсчетом числа растений (или побегов для видов с клональным ростом), «кошением» насекомых сачком, анализом биоты в пробе почвы или воды и т.д. 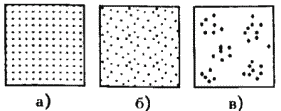 Рис. 13. Типы распределения особей популяции в пространстве: а – регулярное, б – случайное, в – групповое. Выделяют особей популяции в пространстве (рис. 13): – случайное: местонахождение одной особи не зависит от другой. Случайно распределены особи большинства популяций, если местообитания однородны и достаточно благоприятны, а плотность популяции не очень высока; – групповое (контагиозное): этот тип распределения характерен для популяций в мозаичных экосистемах, например в саваннах деревья распределены группами, и соответственно группами распределены обитающие в них популяции птиц и насекомых. Этот же тип распределения отмечается у животных, ведущих групповой образ жизни (сайгак, дзерен) и формирующих колонии (мышевидныех грызунов), а также у клональных растений, разрастающихся пятнами (коротконожки перистой). Групповое размещение особей жертв осложняет хищникам их поиск, может сопровождаться эффектом взаимного благоприятствования, быть средством регулирования температуры тела у животных (см. 4.4.1) и т.д. Таким образом, за наблюдаемым «групповым распределением» могут стоять совершенно разные биологические факторы; – регулярное: расстояние между особями, составляющими популяцию, более или менее одинаковое. Типичным примером является размещение деревьев во фруктовом саду. Однако и среди многих видов птиц, которые разделяют территорию на охотничьи наделы, также возможно распределение, близкое к регулярному. Важным параметром для характеристики популяций животных является величина индивидуального надела (для растений – площади питания). Эти показатели зависят от размера особей: естественно, что для зайца и лося или для дуба и копытня они будут не сопоставимы по размеру. Зависит индивидуальный надел и от уровня обеспеченности ресурсами. Так в национальном парке Найроби в тех районах, где много копытных, индивидуальный надел льва составляет 25–50 км , а там, где продуктивность саванны низка и соответственно мало копытных, – в 10 раз больше. Контрольные вопросы1. Чем отличаются понятия «плотность популяции» и «численность популяции»? 2. Какие факторы определяют размер популяции? 3. Расскажите о типах распределения популяций в пространстве. 4. Что такое «охотничий надел»? 6.5. Гетерогенность популяций Любая природная популяция гетерогенна, т.е. состоит из особей, различающихся по фенотипическим и (или) генотипическим признакам. Одна из форм фенотипической гетерогенности – присутствие в популяции особей разного возраста (разных возрастных когорт). Однако даже в составе одной возрастной когорты могут быть индивидуумы, развитые лучше и хуже, т.е. обладающие разным виталитетом (Злобин, 1993, 1994). Наиболее доступным и информативным показателем виталитета у растений является их размер (вес): чем растение лучше развито, тем выше его виталитет. Фенотипическая дифференциация животных не столь наглядно выражена, как у растений, тем не менее в их популяциях возникают «социальные структуры» – семейные группы, в которых дифференцируются процветающие, средние и слабые особи. Лидеры семейных групп всегда отличаются более мощным сложением, которое позволяет им легче отстаивать свои права на лучшие условия. Как правило, слабые особи оказываются оттесненными к периферии группы и становятся добычей хищников. Фенотипическое разнообразие организмов в популяциях повышает полноту использования ресурсов (даже стадо из коров и телят полнее использует травостой пастбища, чем стадо только из коров или только из телят). В популяциях растений часто отмечается значительная генотипическая вариация за счет сосуществования нескольких экотипов, которые определяются как «…внутривидовые генетически предопределенные локальные соответствия между организмами и средой» (Бигон и др., 1989, т. 1, с. 49). Удивительные примеры сосуществования экотипов клевера ползучего были выявлены Р. Теркингтоном и Дж. Харпером (Turkington, 1978; Turkington, Harper, 1979). Клевер ползучий легко размножается вегетативно, авторы клонировали особи клевера, которые произрастали рядом с разными злаками (ежой сборной, бухарником, райграсом многолетним и др.). Как оказалось, отношения соседства привели к тонкой биотической дифференциации – отбору особых экотипов клевера, которые в культуре «узнавали» своего соседа и отвечали на это усилением роста. В последние годы большой материал о генотипическом разнообразии внутри популяций растений получен методами изоферментного анализа (изоферментных генетических маркеров). В частности, было выявлено, что в большинстве популяций древесных растений отмечается достаточно высокое генотипическое разнообразие, возрастающее в экстремальных условиях у границ экологического ареала вида. Генотипическая вариация в популяциях некоторых видов животных, видимо, ниже, чем в популяциях растений, так как, обладая подвижностью, животные разных экотипов распределяются по популяциям или микросайтам внутри одной популяции. В то же время отмечены случаи сосуществования в одной популяции животных нескольких (чаще двух) экотипов у малоподвижных видов, таких, как улитки (Бигон и др., 1989). В популяциях саранчей есть две формы, резко отличающиеся по внешнему виду – «стационарная» (одиночная) и мигрирующая (стадная), причем соотношение этих форм меняется в зависимости от условий года. Генотипическая гетерогенность популяций, также как и фенотипическая, повышает эффективность использования ресурсов и способствует повышению продуктивности и устойчивости. Например наличие в составе популяции растений раннецветущего и позднецветущего экотипов повышает ее устойчивость к заморозкам, наличие экотипа, более активно накапливающего цианиды – устойчивость к фитофагам и т.д. Гетерогенность природных популяций моделируется в практике сельского хозяйства: используются смеси из нескольких сортов культурных растений с разными экологическими особенностями (более засухоустойчивого и менее засухоустойчивого, высокого и низкого и т.д.). Такие смеси сортов дают более устойчивый урожай, хотя в отдельные годы, наиболее благоприятные для одного или другого сорта-экотипа, его урожай в чистом посеве может быть выше. Наличие экотипов, устойчивых (преадаптированных) к действию гербицидов, объясняет феномен быстрого «приспособления» сорных видов к химическим мерам контроля их популяций. Подобные экотипы могут отбираться и по устойчивости к загрязнению почвы тяжелыми металлами. Генотипическое разнообразие внутри популяций ставит дополнительные задачи перед охраной биоразнообразия, которая должна обеспечивать сохранение не только видов, но и их экотипов. Контрольные вопросы1. Какую роль играет фенотипическая дифференциация особей в популяциях растений и животных? 2. Почему генотипическая дифференциация особей в популяциях растений обычно выше, чем в популяциях подвижных животных? 3. Как используется свойство гетерогенности популяций в сельском хозяйстве? Темы докладов на семинарских занятиях1. Конкуренция как основная форма взаимоотношений особей в популяции. 2. Положительные взамодействия особей в популяциях. 3. Гетерогенность популяций как адаптация для повышения их устойчивости. Глава 7. Динамика популяций Изучение изменений признаков популяций во времени – наиболее сложный раздел популяционной экологии, который включает характеристику закономерностей динамики численности и биомассы популяций. Сложность оценки этих процессов заключается в том, что их результаты интегрально отражают действие множества взаимозависимых факторов. По этой причине даже после того, как описана закономерность изменения размера популяции, далеко не всегда ее можно однозначно объяснить. Главный фактор, который вызвал этот процесс, может влиять на него не только прямо, но и опосредствованно через один или несколько факторов-посредников. При изучении динамики популяций важен учет «биологического времени», на основании которого устанавливается минимальная длительность периода наблюдений. Так для изучения закономерностей динамики популяции тли достаточно нескольких недель, полевки – нескольких лет, а долгожителя-слона – нужны десятки лет. Соответственно для изучения одноклеточных водорослей достаточно нескольких дней, изучение криля требует нескольких месяцев, а наблюдения за популяцией китов, как и слонов, проводятся в течение десятков лет. 7.1. Динамические характеристики популяций Плотность популяции регулируется четырьмя параметрами: 1. рождаемостью – числом особей, родившихся за определенный промежуток времени. Этот промежуток устанавливается в соответствующем масштабе биологического времени. Для бактерий он может быть равен одному часу, для планктонных водорослей – суткам, для насекомых – неделе или месяцу, для крупных млекопитающих (включая человека) – году; 2. смертностью – числом особей, умерших за ту же единицу времени (неважно, своей смертью или погибших, например съеденных); 3. скоростью иммиграции особей – числом особей, появившихся в данной популяции, из других популяций (за ту же единицу времени); 4. скоростью эмиграции особей – числом особей, покинувших данную популяцию за единицу времени. Формула изменения численности популяции выглядит следующим образом:  Если пренебречь показателями иммиграции и эмиграции особей (их, как отмечалось, почти нет у растений), то можно оценить мгновенную скорость роста популяции, т.е. баланс между рождаемостью и численностью за единицу времени. У стабильных популяций мгновенная скорость роста равна нулю, у растущих – является положительным числом, у разрушающихся – отрицательным. Впрочем, даже у стабильных популяций продолжаются процессы циклических изменений численности (см. 12.2). Контрольные вопросы1. Дайте определение четырем основным параметрам, определяющим динамику популяции. 2. Напишите формулу изменения численности популяции. 3. Что такое мгновенная скорость роста популяции? 7.2. Кривые выживания Для изучения закономерностей динамики популяций составляются таблицы выживания. В этих таблицах строками отражаются классы возраста, а в столбцах показывается число особей, которые сохранились или погибли. Величина градаций классов зависит от продолжительности жизни изучаемых организмов (т.е. от биологического времени). Для человека используют интервал в 5 лет, для многих насекомых – одну неделю. Если есть возможность длительное время следить за динамикой вымирания особей в популяциях (регистрировать возраст наступления смерти всех членов одной возрастной когорты, т.е. группы особей, родившихся за короткий относительно общей продолжительности жизни организма период), то составляют динамические таблицы выживания. Однако для долгоживущих или подвижных видов получить данные для построения динамических таблиц крайне трудно. По этой причине составляют таблицы выживания на основании краткосрочных наблюдений за смертностью во всех возрастных группах. Такие таблицы называются статическими, их пример – данные о демографии женской части населения Канады (табл. 6). Таблица 6 Статическая демографическая таблица женского населения Канады на 1980 г. (по Krebs, 1985) 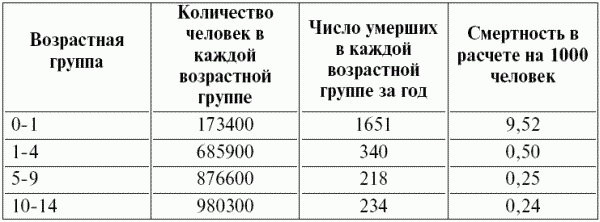 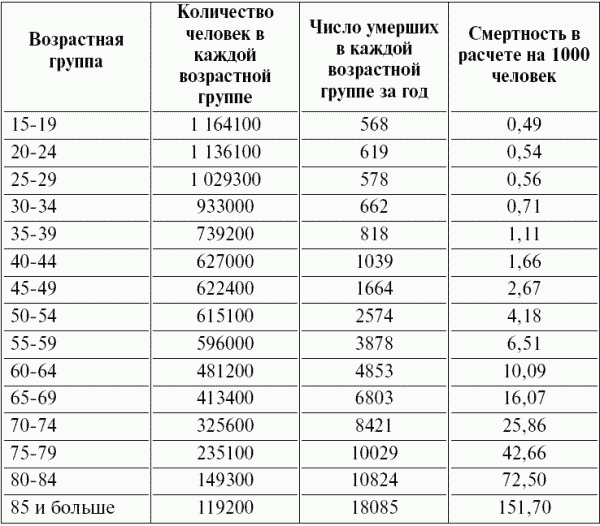 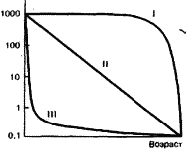 Рис. 14. Три типа кривых выживания. Пояснения в тексте. На основе таблиц выживания строят кривые выживания (рис. 14). Р. Перль предложил различать три типа таких кривых. Кривая I типа (сильно выпуклая) соответствует ситуации, когда смертность ничтожно мала в молодом и среднем возрасте, но в старом возрасте быстро увеличивается и все особи погибают за короткий срок. Перль назвал эту кривую «кривой дрозофилы». К кривой этого типа приближается кривая выживания человека в развитых странах. Кривая II типа (диагональная) представляет ситуацию, когда во всех возрастных классах смертность особей одинакова. Такова динамика популяций многих рыб, пресмыкающихся, птиц, многолетних травянистых растений. Кривая III типа (сильно вогнутая) выражает другой случай – массовую гибель особей в начальный период жизни, а затем низкую смертность выживших особей. Эту кривую Перль назвал «типом устрицы». Ей соответствует и возрастная динамика большинства видов деревьев: высока смертность всходов и молодых растений, однако с возрастом интенсивность самоизреживания резко снижается, и постепенно древостой достигает «конечной плотности», которая отражает особенности биологии вида и условий среды. Она тем ниже, чем благоприятнее условия (выше бонитет насаждений). Так по А.П. Шенникову (1964), «конечная» плотность спелого древостоя в хороших условиях (бонитет I) составляет у ели – 724, у сосны – 470, у дуба – 309 деревьев на 1 га. Однако при худших условиях (бонитет IV–V классов) количество деревьев резко увеличивается и составляет соответственно 2095, 1310, 778. Лесоводы знают эту закономерность и проводят «рубки ухода» как меру по ускорению процесса самоизреживания за счет вырубания более слабых деревьев. Следует заметить, что феномен самоизреживания вследствие конкуренции особей внутри популяции и выживания наиболее сильных характерен только для растений. Как подчеркивают М. Бигон и др. (1989), у животных процесс самоизреживания не происходит, и уменьшение плотности популяций имеет более сложную природу. Контрольные вопросы1. Что такое таблица выживания? 2. Расскажите о кривых выживания. Приведите примеры популяций, соответствующих кривым выживания разного типа. 3. Как зависит конечная плотность спелого древостоя от благоприятности условий среды? 7.3. Модели роста популяций В экологии существует несколько моделей роста популяций (т.е. закономерностей изменения численности популяции при ее росте «от нуля»), главные из них – экспоненциальная и логистическая. О тенденции быстрого неограниченного увеличения числа особей вида знал К. Линней. Однако представления об экспоненциальном росте популяций в начале прошлого столетия сформулировал Т. Мальтус: количество особей в популяции увеличивается в геометрической прогрессии. В качестве примера Мальтус привел гипотетический рост народонаселения страны при условии естественной смертности (при отсутствии эпидемий и войн). Представления о способности любой популяции к экспоненциальному росту является краеугольным камнем популяционной экологии. П.В. Турчин (2002) считает экспоненциальный рост главным законом экологии, близким по значению к закону Ньютона в физике. Модель экспоненциального роста описывается J-образной кривой: в условиях постоянного поступления ресурсов скорость роста популяции увеличивается и кривая взмывает вверх (рис. 15). Модель может быть описана уравнением:
в котором Nt – численность популяции через очередной промежуток времени (t), N0 – исходная численность, е – основание натурального логарифма, r – коэффициент размножения (репродуктивный потенциал, разность относительной рождаемости и относительной смертности, т.е. число родившихся или умерших особей отнесенное к числу особей популяции в начале промежутка времени t). 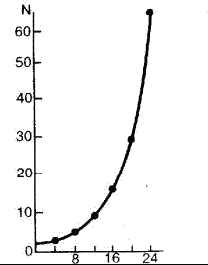 Рис. 15. Экспоненциальная модель роста численности популяции одноклеточного организма, делящегося каждые 4 часа. Чтобы рост популяции соответствовал этой модели, величина коэффициента r должен быть постоянной, т.е. должно быть постоянным среднее количество потомков на одну особь (если r = 0, т.е. рождаемость равна смертности, то численность популяции не растет). В зависимости от величины r увеличение численности особей может быть быстрым и достаточно медленным. Ч. Дарвин рассчитал потенциальные возможности роста популяций разных организмов при реализации экспоненциальной модели. По его оценкам, число потомков одной пары слонов – животных, размножающихся чрезвычайно медленно, – через 750 лет достигнет 19 млн. Если же обратиться к организмам, живущим не так долго и размножающимся более быстро, то цифры будут еще более впечатляющими. У бактерий, которые делятся каждые 20 минут, из одной бактериальной клетки через 36 часов может образоваться биомасса, которая покроет весь земной шар слоем толщиной 30 см, а еще через 2 часа – слоем в 2 м. «Поскольку ни бактерии, ни слоны не покрывают землю сплошным слоем, очевидно, что на самом деле в природе экспоненциальный рост популяций организмов или не происходит вообще, или же происходит, но в течение непродолжительного времени, сменяясь затем спадом численности или выходом ее на стационарный уровень» (Гиляров, 1990, с. 77). В природе экспоненциальный рост численности популяций наблюдается в сравнительно кратковременные периоды их жизни при особо благоприятных условиях, когда постоянно пополняются ресурсы. Так в озерах умеренных широт весной после таяния льда в приповерхностном слое воды содержится много биогенных элементов. По этой причине после прогревания воды здесь наблюдается быстрый рост численности диатомовых и зеленых водорослей. Однако он также быстро прекращается, когда эти ресурсы оказываются израсходоваными и, кроме того, зоопланктон начнет активно выедать водоросли (т.е. за счет регулирования плотности популяций «снизу» и «сверху»). Пример экспоненциального роста популяции – история интродукции северного оленя на острова. Так от 25 особей (4 самца и 21 самка), завезенных в 1911 г. на остров Святого Павла (Берингово море), к 1938 г. сформировалась популяция из 2000 оленей. Однако затем последовал спад численности, и к 1950 г. сохранилось всего 8 особей. Причина краха популяции – нарушение отношений «растение – фитофаг» (см. 8.3) ввиду отсутствия в пищевой цепи третьего звена – хищника. Логистическая модель роста популяций, описываемая S-образной кривой (медленный рост – быстрый рост – медленный рост, рис. 16), была предложена также в начале 19-го столетия бельгийским математиком П.-Ф. Ферхюльстом, а затем уже в 20-е гг. нашего столетия переоткрыта американскими учеными Р. Перлем и Л. Ридом. П.В. Турчин считает эту модель отражением закона «самоограничения роста любой популяции». 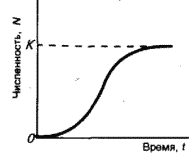 Рис. 16. Логистическая модель роста популяции. К – предельная численность Причины замедления роста популяции могут быть самыми различными: выедание ресурсов, влияние эффекта скученности (у грызунов при этом снижается интенсивность репродуктивного процесса), отравление местообитания прижизненными выделениями, выедание популяции хищниками и т.д. Тем не менее и эта кривая является идеализацией, так как крайне редко проявляется в природе. Очень часто после того, как рост популяции выйдет на плато (достигнет предела К, соответствующего количеству ресурсов), происходит внезапное уменьшение ее численности, а потом популяция вновь быстро растет. Таким образом, ее динамика оказывается состоящей из повторяющихся логистических циклов. Такая циклическая динамика наблюдается, например, в популяциях тундровых леммингов, которые питаются мхами и лишайниками. Они продолжают активную жизнедеятельность под снегом и выедают свою кормовую базу настолько, что прекращают размножаться, а затем начинают умирать от бескормицы. После того, как мхи отрастут, начинается новый подъем численности леммингов. Возможны колебания численности популяции под влиянием погодных условий, паразитов и хищников. Существует особый вариант регулирования плотности популяций, который называется «оппортунистическим», т.е. не укладывающимся в «правильные» законы, описанные экспоненциальной или логистической кривой. У эксплерентов (r-стратегов) происходят вспышки численности в том случае, если появляются обильные ресурсы. При этом рост численности происходит либо за счет того, что начинают развиваться особи из покоящихся диаспор (скажем, почвенного банка семян), либо за счет массового «десанта» в стадии яиц (скажем, мух, налетевших на труп животного). Так как конкуренция в силу обилия ресурсов слаба то израсходовав их изобилие, популяция погибает целиком. У растений-эксплерентов при повышении плотности популяции конкуренция возрастает, но самоизреживания (как у деревьев-виолентов) не происходит, а уменьшается размер особей в десятки и сотни раз. При этом растения проходят весь жизненный цикл и способны дать семена. Дж. Харпер (Harper, 1977) назвал такой тип регуляции плотности популяций растений-однолетников «пластичностью» и противопоставил его самоизреживанию. Эти два типа регулирования плотности в популяциях растений связаны переходом: у большинства видов с вторичными стратегиями при повышении плотности популяции происходит одновременно и уменьшение размера особей, и самоизреживание. На знании этих закономерностей построено обоснование нормы высева культурных растений. Вначале при увеличении нормы высева урожай растет, но потом начинает снижаться (рис. 17). Выбирается та норма высева, которая обеспечивает максимальный урожай. Впрочем, иногда ее несколько завышают, чтобы культурные растения могли подавлять популяции сорных растений. При усилении гербицидного контроля в этом нет необходимости. 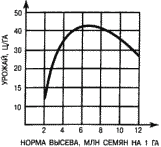 Рис. 17. Зависимость урожая пшеницы от нормы высева при оптимальных экологических условиях. Контрольные вопросы1. Охарактеризуйте экспоненциальную модель роста популяции. 2. Почему модель экспоненциального роста редко наблюдается в природных популяциях? 3. Из каких фаз состоит логистическая модель роста популяции? 4. Какие причины вызывают циклическую динамику популяций? 5. Какие популяции называются оппортунистическими? 7.4. Возрастной состав популяций Кривые выживания могут реализовываться при разном характере динамики популяций: при одновременном «старте» популяций, заселяющих свободное пространство, или при постоянном «популяционном потоке», когда часть особей вымирает, а часть – занимает освободившееся место (ситуация «вокзала», в которой число уезжающих пассажиров постоянно компенсируется вновь прибывающими). В результате в разных популяциях при единовременном учете выявляется разный возрастной состав. Определить абсолютный возраст особи можно не у всех видов. Это несложно сделать для деревьев, воспользовавшись специальным буром, которым извлекают столбик древесины – керн и подсчитывают на нем число годичных колец. У дерева конкретного вида в конкретных условиях (т.е. при одном бонитете) можно определить возраст с точностью до 5 лет по диаметру ствола. У хвойных возраст определяется по числу мутовок побегов на стволе. Однако у трав определение абсолютного возраста затруднительно, и потому оценивают их «возрастное состояние» (стадию). У высших споровых растений (папоротники, хвощи, плауны) выделяются стадии спор, заростков-гаметофитов, молодых спорофитов и взрослых спорофитов. Большой опыт выделения возрастных состояний растений накоплен ботаниками-демографами (Л.Б. Заугольновой, О.В. Смирновой, Л.И. Жуковой и др., табл. 7). Таблица 7 Периодизация онтогенеза цветковых растений (по Жуковой, 1987)  По информативности оценка «возрастного состояния» во многом уступает оценке абсолютного возраста, так как в плохих условиях молодое растение может «выглядеть старым», и, наоборот, при хороших условиях за растение среднего возраста можно принять молодую особь. Соответственно возможны ошибки и при определении возрастного состава популяций и воценке тенденций их динамики. Т.А. Работнов (1992) по соотношению растений разных возрастных групп предложил различать популяции инвазионные (состоят в основном из молодых особей), нормальные (более или менее равномерно представлены все когорты) и регрессивные (преобладают когорты сенильных особей). По возрастному составу популяций растений прогнозируется динамика сообществ и оценивается риск гибели популяций, что особенно важно для редких видов. Исследователи лесов по возрастному составу популяций деревьев (и соответственно отнесению их к инвазионным, нормальным или регрессивным) определяют тенденции «смены пород» в древостое. Если, к примеру, в пойменном тополевом лесу отсутствует возобновление тополя и массово представлены молодые особи вяза, то очевидно, что произойдет смена этого леса вязовым. У организмов с метаморфозом группа особей одинакового «физиологического» возраста представляет одну стадию развития. Так в популяциях насекомых различают стадии яиц, личинок, куколок, взрослых особей. Возрастной состав популяций графически изображается в виде возрастных пирамид. Чаще всего возрастные пирамиды строят для того, чтобы показать демографическую ситуацию в разных популяциях человека. На рис. 18 показаны две пирамиды. Первая характеризует население демографически неблагополучных стран с высокой детской смертностью и низкой продолжительностью жизни (кривая выживания II типа), а вторая – население демографически благополучной страны с низкой рождаемостью, низкой смертностью и высокой продолжительностью жизни (кривая выживания I типа).  Рис. 18. Возрастные пирамиды населения демографически наблагопоучных (а) и демографически благополучны (б) стран, 1985 г. и 2025 г.(прогноз). Контрольные вопросы1. Как строятся пирамиды возрастного состава популяции? 2. Чем отличаются понятия «возраст» и «возрастное состояние»? 3. Приведите примеры возрастных стадий в популяциях растений и насекомых. 4. Расскажите о классификации популяций растений по возрастному составу (по ТА. Работнову). Темы докладов на семинарских занятиях1. Различия динамики популяций животных и растений. 2. Факторы, влияющие на демографические параметры населения страны. 3. Успехи и проблемы изучения возрастного состава популяций растений. Глава 8. Взаимоотношения популяций Взаимодействие популяций (и видов, которые они представляют) – это вопрос, который с равным успехом может рассматриваться в рамках популяционной экологии и при изучении экосистем. С одной стороны, взаимоотношения являются биотическим фактором, влияющим на организм (это и есть влияние «друзей» и «врагов», по определению Э.Геккеля). С другой стороны, взаимодействия организмов – это ячейки системы связей, формирующих экосистему, определяющих распределение ресурсов между организмами одного трофического уровня и закономерности передачи вещества и энергии с одного трофического уровня на другой. 8.1. Классификация взаимоотношений Взаимоотношения организмов разнообразны (табл. 8). Они разделяются на горизонтальные – между организмами одного трофического уровня (как внутри вида, так и между видами) и ветрикальные – между организмами разных трофических уровней. Взаимоотношения первого рода, как правило, носят характер конкуренции, но могут на некоторых этапах жизни организмов быть мутуализмом (т.е. взаимопомощью). Взаимоотношения второго рода более разнообразны: «растение – фитофаг», «хищник – жертва» (иногда эти два типа взаимоотношений объединяют, так как растения по существу тоже жертвы), «паразит – хозяин», мутуализм, комменсализм, аменсализм. Таблица 8 Типы взаимоотношений видов в экосистеме  Примечание. Использованы следующие обозначения: 0 – отсутствие взаимоотношений, плюс – положительное влияние, минус – отрицательное влияние. Кроме материальных взаимоотношений (конкуренции за ресурсы или передачи вещества и энергии при хищничестве или паразитизме), возможны сигнальные (информационные) взаимоотношения. Однако эти сигнальные взаимоотношения тесно переплетены с материальными и лишь корректируют распределение ресурсов между особями или передачу вещества и энергии с одного трофического уровня на другой. Разделение взаимоотношений в естественных экосистемах по «полезности» и «вредности» некорректно: любые взаимоотношения помогают поддерживать экологическое равновесие и в конечном итоге являются «полезными» для всех видов, которые входят в состав экосистемы. О полезности и вредности отношений между организмами можно говорить только в том случае, если в эти отношения вмешивается человек. Понятие «экологическое равновесие» подвергается жесткой критике некоторых экологов, которые считают, что этот феномен не существует в природе, а само понятие уходит корнями в представления Карла Линнея о божественной гармонии природы (см. 1.1). Оппоненты понятия «экологическое равновесие» считают, что любые взаимоотношения организмов и любая экосистема как сумма этих взаимоотношений всегда динамичны, так как испытывают влияние множества факторов. Однако мы полагаем, что понятие экологического равновесия, если оно не абсолютизируется, а рассматривается как некоторая условность (подобная «идеальному газу» или «ускорению» под действием силы земного притяжения), полезно. Системы «хищник – жертва», «паразит – хозяин» и более сложные элементы экосистемы, такие, как пищевые цепи, в состоянии экологического равновесия характеризуются соотношением численности и биомассы взаимодействующих организмов, которые колеблются вокруг некоторой средней величины, соответствующей «экологическому равновесию». Это понятие необходимо при разработке системы рационального использования и охраны природы. Возможно, что более точно этот феномен отражают термины «динамическое равновесие» или «подвижное равновесие». Контрольные вопросы1. Назовите основные типы вертикальных и горизонтальных отношений между видами. 2. Почему некорректно разделять взаимоотношения популяций в природе на «полезные» и «вредные»? 3. Что понимается под экологическим равновесием? 8.2. Конкуренция Конкуренция – это соревнование организмов одного трофического уровня (между растениями, между фитофагами, между хищниками и т.д.) за потребление ресурса, имеющегося в ограниченном количестве. Д.Тилман (Tilman, 1982, 1983) подчеркивает, что особую роль играет конкуренция за потребление ресурсов в критические периоды их дефицита (например между растениями за воду в период засухи или хищниками за жертвы в неблагоприятный год). Конкуренция играет большую роль в определении видового состава экосистем. Принципиальных различий у межвидовой и внутривидовой (внутрипопуляционной) конкуренции нет. Возможны как случаи, когда внутривидовая конкуренция является более острой, чем межвидовая, так и наоборот. При этом интенсивность конкуренции внутри популяции и между популяциями может меняться в различных условиях. Если условия неблагоприятны для одного из видов, то конкуренция между его особями может усиливаться. В этом случае он может быть вытеснен (или чаще – потеснен) видом, для которого эти условия оказались более подходящими. Однако в многовидовых сообществах пар «дуэлянтов» чаще всего не образуется, и конкуренция носит характер диффузной: много видов одновременно конкурируют за один или несколько факторов среды. «Дуэлянтами» могут быть лишь массовые виды растений, которые делят один и тот же ресурс (например, деревья – липа и дуб, сосна и ель и т.д.). У растений возможна конкуренция за свет, за ресурсы почвы и за опылителей. На почвах, богатых ресурсами минерального питания и влагой, формируются густые сомкнутые растительные сообщества, где лимитирующим фактором, за который конкурируют растения, является свет. При дефиците в почве влаги или элементов минерального питания полог растений бывает разомкнутым и они не конкурируют за свет, а соревнуются за потребление почвенных ресурсов (рис. 19).  Рис. 19. Основной комплексный градиент конкуренции растений (по Tilman, 1988). При конкуренции за опылителей побеждает тот вид, который более привлекателен для насекомого. Так одним из факторов быстрого распространения в Европе гималайского заносного вида недотрога железконосная (Impatiens glandulifera) является то, что он продуцирует больше нектара, чем его конкуренты в тех же влажных местообитаниях – чистец болотный, дербенник иволистный. Кроме того, нектар недотроги слаще (Chittka, Schurkens, 2001). У животных конкуренция происходит за ресурсы пищи, например травоядные конкурируют за фитомассу При этом конкурентами крупных копытных могут быть насекомые, подобные саранче, или мышевидные грызуны, способные в годы массового размножения уничтожить большую часть травостоя. Хищники конкурируют за жертвы. Поскольку количество пищи зависит не только от экологических условий, но и от площади, где воспроизводится ресурс, конкуренция за пищу может перерастать в конкуренцию за занимаемое пространство, т.е. быть не только эксплуатационной, но и интерференционной (см. 8.2). Снижение конкуренции в этом случае возможно при разделении территории на «охотничьи наделы» или на «загоны для выпаса». К примеру, косяки – семейные группы башкирской лошади, в поведении которых сохранились черты их диких предков, рассредоточиваются по «загонам для выпаса», что снижает конкуренцию за фитомассу и способствует равномерному использованию травостоев. По этой причине лошади – это идеальные фитофаги для особо охраняемых территорий в степной зоне. Большую роль в разделе территории играют сигнальные взаимоотношения (см. 8.8). Как и в отношениях между особями одной популяции, конкуренция между видами (их популяциями) может быть симметричной или асимметричной. При этом ситуация, когда условия среды одинаково благоприятны для конкурирующих видов, встречается довольно редко, и потому отношения асимметричной конкуренции возникают чаще, чем симметричной. При флюктуирующих ресурсах, что обычно в природе (увлажнение или элементы минерального питания для растений, первичная биологическая продукция для разных видов фитофагов, плотность популяций жертв для хищников), поочередно получают преимущества разные конкурирующие виды. Это также ведет не к конкурентному исключению более слабого, а к сосуществованию видов, которые поочередно попадают в более выгодную и менее выгодную ситуацию. При этом ухудшение условий среды виды могут переживать при снижении уровня метаболизма или даже перехода в состояние покоя. Кроме того, на исход конкуренции часто влияет то, какой из видов первым начал заселять экотоп (принцип лотереи, см. 12.8). Это особенно характерно для маловидовых сообществ водных растений, где более слабый вид может удерживать занятое место, если ему повезло и он его занял первым. Впрочем, лотерея может влиять и на состав сообществ с большим числом претендентов на свободное место. В тропическом лесу на одном гектаре может быть до 150 видов деревьев, и потому занять место выпавшей особи могут представители разных видов. Влияет на исход конкуренции и то, что победить в конкурентной борьбе больше шансов имеет популяция, в составе которой больше особей и которая, соответственно, будет более активно воспроизводить «свою армию» (так называемый масс-эффект). Наконец, конкуренция между видами протекает на фоне отношений с организмами других трофических уровней (хищниками и паразитами). Это также влияет на исход конкуренции, так как более привлекательный как пищевой ресурс вид имеет меньше шансов победить в конкуренции. В итоге в естественных экосистемах виды сосуществуют даже при наличии асимметричной конкуренции, которая должна была бы привести к вытеснению одного из видов. Конкурентное исключение чаще всего наблюдается только в искусственных условиях «микрокосма», когда два конкурирующих вида изолированы и помещены в условия стабильной среды (например в смешанном посеве двух культурных растений с разными конкурентными возможностями). В естественных экосистемах существуют и специальные механизмы, которые снижают конкуренцию. Главный механизм – дифференциация экологических ниш (см. 9), при которой разные организмы используют разные ресурсы. Контрольные вопросы1. Дайте определение конкуренции. 2. Имеются ли принципиальные отличия внутривидовой конкуренции от межвидовой? 3. Какая конкуренция называется асимметричной? 4. Что такое диффузная конкуренция? 5. Приведите примеры конкуренции животных за разные ресурсы? 6. За какие ресурсы среды конкурируют растения? 7. Чем отличаются эксплуатационная и интерференционная виды конкуренции? 8. Как влияет на конкуренцию «принцип лотереи»? 9. Как влияет на конкуренцию «масс-эффект»? 10. Какое значение для конкуренции имеют флюктуации количества ресурсов? 8.3. Взаимоотношения «фитофаг – растение» Взаимоотношения «фитофаг – растение» – это первое звено пищевой цепи, в котором вещество и энергия, накопленные продуцентами, передаются консументам. Для растений в равной мере «невыгодно», чтобы их съели до конца или не съели вовсе. По этой причине в естественных экосистемах проявляется тенденция формирования экологического равновесия между растениями и поедающими их фитофагами. Для этого растения: – защищаются от фитофагов колючками, образуют розеточные формы с прижатыми к земле листьями, малодоступными для пасущихся животных; – защищаются от полного выедания биохимическим путем, продуцируя при усилении поедания токсичные вещества, которые делают их менее привлекательными для фитофагов (это особенно характерно для медленно растущих патиентов). У многих видов при их поедании образование «невкусных» веществ усиливается; – выделяют запахи, отпугивающие фитофагов. Защита от фитофагов требует значительных затрат энергии, и потому во взаимоотношениях «фитофаг – растение» прослеживается трейдофф: чем растение быстрее растет (и соответственно, чем лучше условия для его роста), тем оно лучше поедается, и наоборот, чем растение медленнее растет, тем оно менее привлекательно для фитофагов. Интенсивное отрастание позволяет растениям с высокой поедаемостью сохраняться и даже доминировать в сообществах. В то же время эти средства защиты не обеспечивают полную сохранность растений от фитофагов, так как это повлекло бы за собой ряд нежелательных последствий для самих растений: – несъеденная степная трава превращается в ветошь – войлок, который ухудшает условия жизни растений. Появление обильного войлока ведет к накоплению снега, задержке начала развития растений весной и как итог – к разрушению степной экосистемы. Вместо степных растений (ковылей, типчака) обильно развиваются луговые виды и кустарники. У северной границы степи после этой луговой стадии вообще может восстановиться лес; – «лишние» листья многих видов трав и кустарников делают крону чрезмерно густой, что ухудшает условия для фотосинтеза (затененные листья «паразитируют», т.е. тратят на дыхание больше органического вещества, чем производят его в процессе фотосинтеза). Этот феномен, который исследовал А.А. Любищев, объясняет благоприятное влияние на урожайность посевов присутствия некоторого количества «вредителей»-фитофагов, которые осветляют полог растений. Поедание некоторого количества листьев побегов как бы запрограммировано у культурных растений в «память» о своем диком прошлом; – в саванне уменьшение потребления побегов деревьев веткоядными животными (антилопами, жирафами и др.) приводит к тому, что их кроны смыкаются. В итоге учащаются пожары и деревья не успевают восстанавливаться, саванна перерождается в заросли кустарников. Кроме того, при недостаточном потреблении растений фитофагами не освобождается место для поселения новых поколений растений. Подобным образом регулируется равновесие между популяциями видов фитопланктона и зоопланктона. Активно поедаемые водоросли быстро размножаются. Некоторые водоросли, напротив, защищаются от выедания специальными выростами на твердых панцирях (как диатомовые) или объединением в большие колонии, которые не могут быть отфильтрованы рачками. Колониальными формами представлено большинство видов цианобактерий. Защите водорослей от выедания помогает их способность образовывать покоящиеся стадии, играющие ту же роль, что и банки семян у растений. Наконец, некоторые водоросли заглатываются планктонными животными-фитофагами, но не перевариваются и выделяются с экскрементами живыми. «Несовершенство» отношений «фитофаг – растение» приводит к тому, что достаточно часто случаются кратковременные вспышки плотности популяций фитофагов и временное угнетение популяций растений, вслед за которыми следует и снижение плотности популяций фитофагов. Например в степях Монголии один раз в 5-7 лет наблюдаются вспышки численности популяций полевки Брандта, которая полностью выедает надземные части степных растений и разрывает дернину. В итоге угнетаются популяции ковылей, а корневищные злаки быстро разрастаются за счет банка вегетативных зачатков (почек на корневищах). Вслед за вспышкой численности наступает депрессия популяций грызунов из-за массового заболевания, а популяции ковылей восстанавливаются. Для обеспечения экологического равновесия в паре «фитофаг – растение» адаптаций растений и животных, как правило, недостаточно. Оно возможно только в случае, если имеется зоофаг, который контролирует плотность фитофага. По этой причине невозможно экологическое равновесие, если из экосистемы исчез хищник или в нее внедрился вид-фитофаг, для которого нет контролирующего его хищника. При разведении сельскохозяйственных животных плотность популяций фитофагов регулируется человеком, который сам устанавливает норму потребления фитомассы животными. В большинстве случаев при этом норма оказывается завышенной, что ведет к развитию процессов пастбищной дегрессии в экосистемах (см. 12.7). Контрольные вопросы1. Расскажите об адаптациях, которые позволяют растениям защищаться от фитофагов? 2. Как фитофаги преодолевают «оборону» растений? 3. Почему для устойчивого экологического равновесия в звене пищевой цепи «растение – фитофаг» необходим хищник или паразит? 8.4. Взаимоотношения «хищник – жертва» Взаимоотношения «хищник – жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка. Как и при отношениях «растения – фитофаг», ситуации, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами, исключающими полное истребление жертв. Так жертвы могут: – убегать от хищника. В этом случае в результате адаптации повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей («принцип Тома и Джерри»); – приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий (например красный цвет, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в листве, а зимой – на фоне белого снега; – распространяться группами, что делает их поиск и промысел для хищника более энергоемким; – прятаться в укрытия; – переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.). В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки). В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.). Полной защиты от хищников всех особей в популяциях животных-жертв также не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге – к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения их численности подрывается кормовая база. И.В. Стебаев и др. (1993) образно сравнивают отношения видов-хищников и их жертв с отношениями между двумя флотами, каждый из которых постоянно усиливает свою огневую мощь и наращивает толщину брони. Но эти флоты никогда не вступают в генеральное сражение. Тем не менее даже при столь богатом арсенале средств защиты у жертв и нападения у хищников экологическое равновесие в паре «хищник – жертва» только за счет их биотических потенциалов невозможно. И в этом случае, как и в отношениях фитофагов и растений, необходимо третье звено пищевой цепи – естественный «враг» хищника – хищник более высокого порядка или паразит. В зависимости от характера жертвы и типа хищника (истинный, убивающий жертву сразу, или пастбищник, использующий ее многократно, как, например, оводы или слепни) возможна разное соотношение динамик их популяций. При этом картина осложняется тем, что хищники редко бывают монофагами (т.е. питающимися одним видом жертвы по типу «преследования»). Чаще всего, когда истощается популяция одного вида жертвы и ее добыча требует слишком больших затрат сил, хищники переключаются на другие виды жертв. Кроме того, одну популяцию жертв может эксплуатировать несколько видов хищников и паразитов. По этой причине часто описываемый в экологической литературе эффект зависимости численности популяций жертв и хищников – пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки – Вольтерры»), – выражен далеко не во всех парах «хищник – жертва». П.В. Турчин (2002) считает осцилляторный (колебательный) характер динамики системы «ресурс – потребитель», которая включает отношения типа «фитофаг – растение» и «хищник – жертва», третьим законом популяционной экологии. При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага. При этом он должен руководствоваться экологическими нормативами «макисмально допустимого урожая, МДУ», отражающими скорость восстановления плотности популяций. К примеру, без риска разрушить популяцию северного оленя из нее ежегодно можно изымать до 40% животных, а из популяции лосей – только 20%. Примерно также показатель МДУ используется при эксплуатации популяций растений. Контрольные вопросы1. Как жертвы «обороняются» от хищников? 2. Как хищники совершенствуют систему преследования жертв? 3. Какие дополнительные условия необходимы для формирования экологического равновесия в паре «жертва – хищник». 4. Почему «эффект Лотки–Вольтерры» в природе проявляется не всегда? 5. Что такое МДУ? 8.5. Взаимоотношения «паразит – хозяин» Паразиты – это организмы, которые питаются за счет организма-хозяина. Это очень разнообразная группа организмов (животные, растения, грибы, бактерии), которую изучает специальная наука – паразитология. Паразиты не убивают хозяина, а длительное время используют его как пищевой ресурс и убежище, лишь сокращая длительность жизни и плодовитость «организма-дома». Паразиты близки к хищникам-пастбищникам, но отличаются тем, что если хищник-пастбищник использует несколько жертв, то паразит связан с одним организмом-хозяином (а если с несколькими, то меняет их в ходе жизненного цикла). Различаются следующие группы паразитов. Биотрофы и некротрофы. Биотрофы всю жизнь питаются за счет живого хозяина, а некротрофы губят его (или часть его тела, например лист растения) и завершают свою биографию как рассматриваемые далее детритофаги. Микропаразиты и макропаразиты. Различаются по размерам. К микропаразитам относятся вирусы, бактерии, микроскопические грибы и простейшие, к макропаразитам – все прочие. Истинные паразиты и паразитоиды. Истинные паразиты всю свою жизнь питаются за счет организма-хозяина (или нескольких хозяев, если в течение жизненного цикла переселяются из организма одного вида в другой). Паразитоиды (как правило, насекомые) на определенных стадиях жизненного цикла ведут свободный образ жизни (питаются как фитофаги или зоофаги). После этого они откладывают яйца в тело организма-хозяина, в котором паразитируют личинки. Паразитоиды представляют переход к хищникам. Насекомые-паразитоиды используются для биологического метода контроля насекомых-вредителей в сельском хозяйстве (трихограмма, теленомус-наездник и др.). Эндотрофные и эктотрофные паразиты. Эндотрофные паразиты живут внутри организма-хозяина (глисты в кишечнике позвоночных, стеблевые нематоды в стеблях хлебных злаков), а эктотрофные – на поверхности организма-хозяина (блохи, вши, клещи и др.). Особый случай эктотрофного паразитизма – образ жизни карликовых самцов глубоководных удильщиков, которые внедряются острыми зубами в тело самки, после чего тела хозяина и паразита сливаются воедино (объединяются даже кровеносные системы). Тело самки в 13 раз больше тела самца. Экологический смысл этой адаптации заключается в повышении вероятности оплодотворения. Удильщики живут в полной темноте, и иные варианты поиска «спутника жизни» реализовать крайне сложно. Суперпаразиты – «паразиты паразитов». Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), верхний уровень представлен бактериями или вирусами. Эту матрешку очень точно описал Дж. Свифт: Под микроскопом он открыл, что на блохе В этом случае из паразитов формируется пищевая цепь. Различаются группы паразитов и по сложности жизненного цикла. Одни виды паразитов передаются при непосредственном контакте особей хозяина (например вирусы и бактерии, вызывающие болезни человека). Другие паразиты перед заражением основного хозяина, в теле которого они образуют потомство, проходят через один или несколько видов промежуточных хозяев (например широкий лентец – паразит человека, но в течение жизненного цикла он проходит через стадии жизни в рачках-циклопах и рыбах). И, наконец, кроме паразитов, существуют еще и полупаразиты. Значительное число видов растений (в средней полосе в первую очередь из семейства норичниковых) сочетает автотрофное питание и паразитирование на корнях растений-хозяев. При этом если каждый вид растений-паразитов (из семейств заразиховых, повиликовых и др.) имеет своего «персонального» хозяина, то число видов-хозяев для одного вида полупаразита исчисляется десятками и сотнями. В этом случае нет необходимости в тонком подстраивании метаболизма хозяина и полупаразита, как при абсолютном паразитизме, полупаразит получает от растения-хозяина только неспецифические элементы питания. Среди полупаразитов есть виды, наносящие вред хозяйству человека. Например погремки (Rhinanthus) при доминировании в луговых сообществах становятся основными продуцентами органического вещества. В этом случае за счет потери энергии при переходе с одного трофического уровня на другой (см. 10.4) биологическая продукция травостоя снижается в 2-3 раза. Причем погремок не поедается сельскохозяйственными животными ни на пастбище, ни в сене. В естественных экосистемах взаимоотношения «паразит – хозяин» являются одним из важных факторов поддержания экологического равновесия. Особенно велика их роль при контроле плотности популяций крупных животных, у которых нет естественных врагов-хищников (слон, бегемот, крокодил, лев и др.). При отсутствии паразитов их отношения с жертвами могли бы быть нарушены. В процессе длительной коэволюции (взаимоприспособления) паразитов и хозяев вырабатываются специальные механизмы, которые позволяют им устойчиво сосуществовать. Защитные реакции хозяев могут быть следующими: – иммунный ответ организма, т.е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов; – сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги; – выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при паразитировании тли); – изоляция органов поражения как «зеленых островов» (формирование галлов у дуба, орешника и других растений после того, как насекомое-паразитоид отложит в ткани листа яйцо); – уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, которые таким образом снижают долю зараженных особей в популяции; – формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям. В естественных экосистемах формирование экологического равновесия между популяциями паразитов и их хозяев – нормальное явление. При этом в отличие от отношений «фитофаг – растение» или «хищник – жертва» оно возможно без третьего участника. В силу того, что паразиты связаны с ограниченным кругом хозяев, эта связь математически описывается проще, чем связь между хищниками и их жертвами. Во многих случаях проявляется модель Лотки–Вольтерры: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев. Ситуация изменяется в антропогенных экосистемах, особенно в сельскохозяйственных, где заражение скота паразитами может привести к гибели многих животных. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим – лямблией), болезнями бактериальной и вирусной природы. Катастрофическими бывают последствия заноса паразитов в новые районы, где у их потенциальных хозяев отсутствуют механизмы защиты от паразитов. В ХХ в. произошли ботанические катастрофы в Америке (гибель зубчатого каштана Castanea dentata от занесенного туда из Китая паразитического гриба Endothia parasitica, вызывающего «рак каштана») и Европе, где от «голландской болезни» почти полностью исчез вяз. Болезнь вызывает гриб Ophiostoma ulmi, который переносится жуком-короедом. К еще более серьезным последствиям привело распространение вируса «коровьей чумы» в саваннах Африки (McNaughton, 1992). Под влиянием этого вируса, который первоначально вызвал болезни у домашнего скота, а затем поразил многие виды диких крупных копытных животных, резко снизилась нагрузка фитофагов на растительность саванны, и в первую очередь на ее древесно-кустарниковый компонент и на его отношения с травяным покровом саванны. Кустарники и деревья стали бурно разрастаться, подавляя травы, что увеличило количество пожаров, которые при таком сомкнутом древесном пологе стали более частыми. После пожаров деревья отрастали плохо и замещались кустарниками, корневища которых позволяли им сохраняться во время пожара. Только в 70-е годы, когда удалось снять влияние на экосистемы вируса «коровьей чумы», вылечив от болезни домашний скот, поставлявший паразита популяциям диких животных через выделение слюны на пастбищах, где их выпас чередовался, процесс был остановлен. Восстановилась плотность популяций животных, питающихся ветками, и соответственно восстанавился баланс между древесно-кустарниковым и травяным компонентами экосистемы саванны, которая приобрела первозданный облик. Есть примеры более сложных взаимоотношений «паразит – хозяин» с посредником. Так гетеротрофное растение-паразит подъельник паразитирует на грибах, разлагающих мертвое органическое вещество, но, кроме того, по гифам микоризного гриба, как по шлангу, выкачивает питательные элементы из корней ели. Контрольные вопросы1. Чем паразиты отличаются от хищников? 2. Расскажите о разнообразии паразитов. 3. Какие защитные реакции против паразитов вырабатываются у хозяев? 4. Расскажите о нарушении экологического равновесия в паре «паразит – хозяин» при вмешательстве человека. 8.6. Мутуализм Мутуализм — это форма взаимоотношений организмов, при которых партнеры получают пользу. Отношениями мутуализма связаны организмы, не конкурирующие за ресурсы. Мутуализм включает разнообразные формы сотрудничества – от облигатного (симметричного или асимметричного), при нарушении которого гибнут оба или один сотрудничающий партнер, до факультативного, которое помогает выживать партнерам, но не является для них обязательным (так называемаяпротокооперация). Рассмотрим основные варианты мутуализма. Растения и микоризные грибы. Такие взаимоотношения с грибами (микотрофия) свойственны большинству видов наземных сосудистых растений (цветковых, голосеменных, папоротников, хвощей, плаунов), что во многом облегчило заселение растениями суши (Заварзин, 2000). Микоризные грибы могут оплетать корень растения и проникать в ткани корня, не нанося ему при этом существенного ущерба (эндотрофные и эктотрофные микоризы). Грибы, не способные к фотосинтезу, получают из корней растений органические вещества, а у растений за счет разветвленных грибных нитей в сотни и тысячи раз увеличивается всасывающая поверхность корней. Кроме того, некоторые микоризные грибы не просто пассивно всасывают элементы питания из почвенного раствора, но и одновременно выступают в роли редуцентов и разрушают сложные вещества до более простых. Кроме того, микоризные грибы, выделяя антибиотики, защищают корни растений от патогенов. Микоризные грибы – «дорогое удовольствие» для растений, так как использование их в качестве посредников для обеспечения элементами питания и водой сопряжено со значительными затратами продуктов фотосинтеза (1/3 или даже 1/2 валовой первичной продукции). По этой причине при улучшении условий минерального питания, например при удобрении лугов, даже типичные микотрофные растения отказываются от микориз и переходят на «самообслуживание». Не тратятся на содержание микориз виды-нитрофилы (распространенные на почвах с высоким содержанием нитратного азота) из семейств маревых, крестоцветных и некоторых других, которые селятся на первых стадиях восстановления экосистем после нарушений (см. 12.6), когда за счет минерализации органического вещества в почве резко возрастает количество нитратов. При этом микоризы, которыми обладают виды следующих стадий сукцессии, выделяют вещества, подавляющие «самостоятельные» растения. Это ускоряет процесс вытеснения нитрофилов. Микоризы нет у водных растений, и сравнительно редко она встречается у растений экстремальных условий – пустынь, горных и арктических тундр. Как подчеркивает Т.А. Работнов (1992), большинство микотрофов – это мезофиты умеренно богатых почв. Микоризы у травянистых растений, как правило, не видоспецифичны (т.е. один вид грибов может формировать микоризу у разных растений), а у древесных – видоспецифичны. Таким образом, плодовые тела подберезовика, подосиновика, масленка или рыжика образуются за счет продукции фотосинтеза соответствующих видов деревьев. Поскольку микоризные грибы оплетают корни нескольких рядом произрастающих растений, по ним возможен горизонтальный перенос элементов питания от одного растения к другому по «гифопроводам». А.М. Гиляров (2003) рассматривает это как «экзаптации на уровне сообществ», т.е. как побочный эффект адаптации микоризного гриба к нескольким видам растений. Данных о количестве веществ, перекачиваемых по микоризам из одного растения в другое, мало. Можно полагать, что оно невелико, тем не менее смягчает отношения конкуренции и повышает общую устойчивость экосистем. Растения и микроорганизмы-азотфиксаторы. Возможны две формы такого мутуализма – облигатный мутуализм и протокооперация. В первом случае азотфиксирующие микроорганизмы живут в корнях растений (бобовых, облепихи, ольхи и некоторых других), вызывая образование клубеньков. Процесс связывания атмосферного азота облигатными азотфиксаторами называется симбиотической азотфиксацией. При протокооперации азотфиксирующие микроорганизмы населяют примыкающую к корням часть почвы (ризосферу) и усваивают органические вещества, которые, как в проточном культиваторе, постоянно выделяются корнями. Такая азотфиксация называется ассоциативной. В целом ассоциативная азотфиксация преобладает в естественных экосистемах, симбиотическая – в агроэкосистемах. Симбиотические микроорганизмы могут жить и в листьях, пример – водный папоротник азолла, распространенный в тропическом поясе. Связанная с азоллой цианобактерия анабена способна за год фиксировать до 1000 кг/га азота (что является бесспорным рекордом, достойным книги Гиннеса). Для сравнения посев клевера в средней полосе способен за год фиксировать до 200 кг/га азота, а люцерны в жарких районах с удлиненным полевым периодом и при поливе – до 700 кг/га (к слову, оптимальная доза внесения азотных удобрений в разных условиях и для разных культур колеблется в пределах 50-200 кг/га; в настоящее время в России в почву в среднем вносится 10 кг/га азотных удобрений в действующем веществе). Обеспечение новых («мертвых») субстратов азотом является необходимым условием для их зарастания. В теплом климате азот в субстрате накапливается в результате симбиотической азотфиксации: пионерами заселения лавовых потоков, отложений речного аллювия, горных осыпей являются бобовые растения (особенно часто из рода люпин). В более прохладном климате азот поставляется в результате ассоциативной азотфиксации: новые субстраты зарастают злаками и осоками. В самых суровых условиях Севера пионерами оказываются цианобактерии, которые обладают уникальной способностью и к фотосинтезу, и к азотфиксации. Мутуалистические взаимоотношения с азотфиксаторами, также как и содержание микориз, обходятся растениям очень дорого: на них затрачивается значительное количество продуктов фотосинтеза (около 1/3). Большими затратами органического вещества на симбиотическую азотфиксацию объясняются более низкие урожаи зернобобовых культур по сравнению со злаками. Тем не менее на биологическую азотфиксацию экологи возлагают большие надежды, она должна во многом заменить техногенную азотфиксацию промышленных предприятий, при которой на производство минеральных азотных удобрений затрачивается очень много энергии. Кроме того, экологически грязным является не только само производство удобрений, но и их использование: при внесении азотных удобрений на поля до 50% их вымывается в окружающую среду, вызывая ее загрязнение (в первую очередь эвтрофикацию водных экосистем, см. 12.7). Растения и насекомые-опылители. Насекомые, переносящие пыльцу, питаются нектаром или пыльцой. Отмечены случаи участия насекомых в опылении даже таких типично ветроопыляемых растений, как злаки. Насекомые-опылители переносят пыльцу с одного цветка на другой на большие расстояния, чем ветер. Если пыльца деревьев за время, пока рыльцевая поверхность сохраняет способность ее воспринимать, может быть перенесена ветром не более чем на 70 м (у трав – менее 10 м), то за это время шмели переносят пыльцу на расстояние до 3 км. Радиус переноса пыльцы пчелами обычно ограничен 1 км. Существует два основных направления развития мутуализма растений и насекомых: узкая и широкая специализация (т.е. в направлении облигатного мутуализма и протокооперации). При узкой специализации эволюция ведет к ограничению числа опылителей: происходит усложнение строения цветка (как у бобовых или губоцветных) таким образом, что нектар становится доступным только для насекомых с определенным типом строения (в первую очередь ротового аппарата). Высшее достижение этого варианта эволюции – взаимоотношения опылителей и некоторых представителей орхидных, которые привлекают самцов насекомых-опылителей, имитируя облик и половые феромоны самок. При широкой специализации спектр опылителей возрастает. Широкий спектр опылителей имеют представители семейства сложноцветных. Этим объясняется их высокая устойчивость в антропогенно нарушенных экосистемах, в которых обеднен состав видов опылителей. По этой причине в современном нарушаемом человеком мире обязательный мутуализм растений и насекомых менее выгоден для обоих партнеров, чем протокооперация. Эффективность протокооперации возрастает по причине неодновременного цветения разных видов, опыляемых одним видом насекомых. Более того, как правило, насекомые посещают цветки именно в апогей их цветения, когда продукция нектара максимальна. (Пчеловоды прекрасно знают о том, что их подопечные сначала посещают один вид растения и только после того, как его цветки минуют пик нектарообразования, переключаются на сбор нектара с цветков другого вида.) В тропиках опылителями некоторых растений являются птицы и летучие мыши. Растения и животные, распространяющие их семена. Распространение плодов (и семян) растений с помощью животных (зоохория) широко представлено в природе. Агентами-распространителями являются птицы, поедающие сочные плоды, медведи, копытные, насекомые. При прохождении через пищеварительный тракт животных семена зоохорных растений не только не перевариваются, но даже повышают всхожесть. Кроме плотных покровов, защищающих семена от переваривания, существуют другие приспособления для зоохории. Так на семенах растений (многих губоцветных, лилейных, маковых, молочайных, лютиковых, сложноцветных), распространяемых муравьями имеются специальные придатки, богатые маслом, которые привлекают муравьев и используются ими в пищу Сухие зоохорные плоды снабжены различными крючочками и щетинками для прикрепления к шерстному покрову животных, например у репейничка, череды, чернокорня, лопуха. С помощью животных распространяются споры некоторых видов грибов и мхов. Водоросли и грибы в лишайнике. В этом случае мутуализм столь обязателен и функции сотрудничающих организмов столь прилажены друг к другу, что по-существу возникает новый «организм второго порядка». Водоросль обеспечивает гриб органическими веществами, гриб водоросль – водой и минеральными элементами. Этот вариант обязательного мутуализма представлен весьма широко: имеются сотни видов лишайников. Лишайники первыми заселяют поверхность скал и широко распространены на Севере в условиях крайней ограниченности ресурсов тепла и элементов минерального питания. Долгие годы в литературе ведется дискуссия о симметричности отношений гриба и водоросли в лишайнике. В последнее время все чаще эти отношения рассматриваются как асимметричные: большую «выгоду» от симбиоза получает гриб («водоросль – это царевна, плененная жестоким драконом»). Млекопитающие и микроорганизмы, населяющие их пищеварительный тракт. Большинство животных, включая человека, но особенно травоядные, сами не в состоянии переваривать пищу, так как не имеют ферментов, разрушающих целлюлозу, и эту роль играют микроорганизмы – бактерии и некоторые простейшие, которые живут в их желудочно-кишечном тракте. В кишечном тракте гладких китов среди 1000 видов бактерий были найдены даже те, которые могут разрушать органические вещества, присутствующие в нефтепродуктах. Возможно, что наличием этих симбионтов объясняется сравнительно высокая устойчивость этого вида китов к нефтяному загрязнению океана. Хемоавтотрофные бактерии и низшие животные. Червеобразные животные-вестиментиферы (тип погонофоры) в стадии личинки являются типичными гетеротрофами со ртом, пищеварительным каналом и анусом. Однако после того как они заглатывают серобактерии, происходит редукция органов пищеварения, клетки животного заполняются серобактериями, и вестиментиферы становятся «симбиотическими автотрофами». В результате мутуализма бактерии получают сероводород, а животное – органическое вещество. Содержание сероводорода в среде, где обитают вестиментиферы, таково, что может погибнуть любой другой организм. Их спасает особый тип гемоглобина, который связывает не только кислород, но и серу. Мутуализм позволяет вестиментиферам очень быстро расти и достигать длины 2,5 м. В.В.Малахов (2001) считает, что мутуализм вестиментифер и бактерий возник в результате развития пищевых отношений типа «жертва – хищник». В гидротермальных оазисах океана и сейчас существуют свободноживущие серобактерии, которые формируют «маты», служащие пищей для многих животных. Начав с питания такими свободноживущими бактериями, вестиментиферы со временем вступили с ними в отношения мутуализма. Подобным образом питаются и другие погонофоры, связанные отношениями мутуализма уже не с серобактериями, а с метанобактериями. Бактерии используют метан, образующийся в нефтяных пластах и поступающий в океан по трещинам в плитах литосферы. Это позволяет использовать погонофор в качестве биологических индикаторов месторождений нефти. Кишечнополостные и водоросли. Водоросли поселяются в теле кораллов, заключенных в известковый скелет, и снабжают животное органическим веществом. Животное поставляет водорослям питательные элементы и дает убежище. В отличие от погонофор кораллы являются гетеротрофами, которые питаются зоопланктоном. Водоросли лишь помогают наиболее эффективно использовать вещества, полученные при гетеротрофном питании. Этим мутуализмом объясняется быстрый рост коралловых рифов. Человек и сельскохозяйственные животные и культурные растения. Этот вариант мутуализма является протокооперацией, тем не менее ни человек при современной плотности населения на планете не может обойтись без сельскохозяйственных животных и растений, ни корова, пшеница или рис не могут выжить без человека. Причиной мутуализма является искусственный отбор, в результате которого из «эгоистических» побуждений человек усиливал у растений и животных эксплерентность и снижал патиентность и виолентность, что лишило эти организмы способности жить без его опеки. Особенно активно это направление селекции развивалось в 60–70-е гг. ХХ в., когда в странах субтропического и тропического поясов произошла Зеленая революция (см. 11.5). Широко распространен мутуализм водорослей и простейших в океанических экосистемах. Некоторые простейшие после поедания водорослей используют их хлоропласты, причем они продолжают работать в теле простейшего до тех пор, пока не износятся, после чего перевариваются. Как протокооперация (хотя и весьма слабая) могут рассматриваться взаимоотношения между бобовыми и злаками в сеяных травостоях: бобовые за счет связи с симбиотическими азотфиксирующими бактериями улучшают условия обеспечения злаков азотом, а вертикально ориентированные листья злаков, пронзающие густой травостой бобовых, понижают уровень конкуренции за свет. Известно множество других «экзотических» вариантов мутуализма: – разведение грибов муравьями и жуками; – отношения африканской птицы медоуказчика и капского медоеда (птица находит пчелиное гнездо, а медоед вскрывает его); – отношения чистильщиков (птиц, рыб) и их «клиентов»: – отношения муравьев и акаций (муравьи охраняют акацию, соком которой питаются, от других фитофагов) и мн. др. Контрольные вопросы1. Какую роль в жизни растений играют микоризные грибы? 2. Какое количество продуктов фотосинтеза затрачивает растение на «содержание» микоризного гриба? 3. В каких случаях растения могут обходиться без микоризы? 4. Расскажите о симбиотической и ассоциативной азотфиксации. 5. В каких экосистемах преобладает симбиотическая азотфиксация? 6. Какую роль играет биологическая азотфиксация в экологизации сельского хозяйства? 7. Дайте оценку роли мутуализма и протокооперации в отношениях растений с насекомыми опылителями. 8. Расскажите о роли зоохории в расселении растений. 9. Какую пользу получают гриб и водоросль от соместеного существования в составе лишайника? 10. Расскажите о мутуализме млекопитающих и микроорганизмов, населяющих их пищеварительный тракт. 11. Рассмотрите вестиментиферы как пример мутуализма животных и хемотрофных бактерий. 12. Какую роль в жизни кораллов играют связанные с ними водоросли? 13. Почему отношения человека и сельскохозяйственных растений и животных рассматриваются как мутуалистические? 8.7. Комменсализм и аменсализм Мутуализм связан плавным переходом с другим вариантом отношений организмов – комменсализмом, при котором сотрудничество выгодно только одному из партнеров. При этом на разных стадиях сотрудничества (или чаще в разных условиях) большую выгоду может получать то один, то другой партнер. Рассмотрим некоторые наиболее характерные случаи комменсализма. «Растения-няни» и их подопечные. Береза или ольха могут быть «няней» для ели: «няня» защищает всходы ели от прямых солнечных лучей, так как на открытом месте елочки вырасти не смогут. В таких же отношениях состоят кустарники из семейств губоцветных и сложноцветных с южноамериканскими кактусами. Кактусы для снижения дневного испарения воды фотосинтезируют при закрытых устьицах (они используют углерод кислот, образованных из захваченного устьицами за ночь углекислого газа) и потому не «потеют». Взрослые кактусы имеют большую фитомассу, и им перегрев не страшен, а молодые могут развиваться только в тени засухоустойчивых кустарников, так как на открытом солнечном свете они гибнут. Весьма характерно, что в более благоприятных климатических условиях (например при подъеме в горы, где климат более прохладный) кактусы не нуждаются в «растениях-нянях». Эпифиты и деревья. Как комменсализм можно рассматривать отношения эпифитов и деревьев-хозяев: они выгодны для эпифита, но безразличны для хозяев, так как эпифиты используют лишь отмершие ткани дерева или скопления пыли опавших листьев. Группа эпифитов достаточно разнообразна и включает лишайники, водоросли, папоротниковидные, цветковые растения. При этом сосудистые эпифиты распространены почти исключительно в теплом тропическом и субтропическом климате. В умеренном климате эпифиты представлены только лишайниками, которые селятся на стволах деревьев, и реже мхами. Отсутствие эпифитов в лесах умеренной полосы связано в первую очередь с их неспособностью переносить холодные периоды года. Как правило, эпифиты – чистые комменсалы, но есть случаи, когда установлено их отрицательное влияние на дерево-хозяина. При этом отрицательное влияние оказывает не сам эпифит, а его микоризный гриб, который ведет себя как паразит. Если эпифитов очень много, то они создают дополнительную нагрузку на ветви, что может вызвать их поломку. Таким образом, «безвредность» эпифитов для деревев относительна. Детритофагия. На те популяции, организмы которых поедаются в мертвом состоянии, детритофаги практически не влияют. Хотя это положение нельзя абсолютизировать: участвуя совместно с редуцентами в процессе минерализации органического вещества, детритофаги способствуют его возврату в окружающую среду и тем самым росту растений и всех организмов, которые связаны с ними в пищевых цепях. В почве сконцентрировано огромное разнообразие детритофагов: число их видов на 1 м 2 почвы в лесу умеренной полосы может превышать 1000, а общая плотность популяций простейших и нематод – 10 млн, ногохвосток (Collembola) и почвенных клещей (Acari) – 100 тыс. и т.д. Почвенные детритофаги разделяются на группы микрофауны (в нее входят простейшие, нематоды и коловратки, а также специализированные животные, питающиеся микроорганизмами, то есть, строго говоря, к детритофагам не относящиеся), мезофауны (животные с шириной тела от 100 мкм до 2 мм), макрофауны (ширина тела от 2 до 20 мм) и мегафауны (ширина тела более 20 мм). В последнюю группу входят мокрицы (Isopoda), многоножки (Diplopoda), дождевые черви (Megadrili), улитки и слизни (Mollusca), личинки некоторых мух (Diptera) и жуков (Coleoptera). Ч. Дарвин считал, что только дождевые черви перерабатывают такое количество растительного детрита, что ежегодное поступление в почву их экскрементов достигает 50 т/га. В тропических районах это количество может быть в три раза выше. Именно благодаря этой армаде детритофагов из мертвого органического вещества (в травяных экосистемах в первую очередь корней растений, в лесных – опада) формируется почва. При этом многие детритофаги одновременно являются и хищниками, так как питаются «бутербродами» из мертвого вещества и содержащихся в нем живых бактерий. Особая группа детритофагов – копрофаги, питающиеся экскрементами. Большинство копрофагов питается экскрементами травоядных животных, которые усваивают поедаемую растительную массу не более чем на 10%, остальное выделяется или в окружающую среду в непереваренном состоянии (существуют даже варианты детритофагии в сельском хозяйстве при повторном скармливании животным специально подготовленных экскрементов). Фауна копрофагов очень разнообразна, особенно в тропиках (в частности в ее составе есть навозный жук Heliocopris dilloni, имеющий размер до 6 см!). Копрофаги часто специализированы для «переработки» определенного типа экскрементов, что создало проблемы после появления в Австралии крупного рогатого скота, навоз которого местные копрофаги, привыкшие к «вкусу» помета кенгуру, перерабатывать отказались. Пришлось отправлять в Австралию жуков-копрофагов из Африки! Количество копрофагов, питающихся экскрементами хищников, много меньше, так как в этом случае корм переваривается более полно, и потому потенциальная пища детритофагов менее питательна. Крупные детритофаги, питающиеся трупами животных, называются мусорщиками (это – ворон, коршун, гриф, шакал, росомаха, барсук, песец и др.). К этой же группе относятся некоторые крупные беспозвоночные, например жук-мертвоед некрофорус, который способен (вдвоем с самкой) закопать труп мыши на глубину до 20 см и там «скормить» его своим личинкам. Среди детритофагов водных экосистем по способу добывания и переработки пищи различают размельчителей, собирателей, соскребателей, фильтраторов. В некоторых случаях возможны отношения типа аменсализм – вредные для одного партнера и нейтральные для другого. Аменсализм представлен в природе как крайний вариант асимметричной конкуренции, когда один вид перехватывает ресурсы у другого, но этот другой столь слаб, что практически не может воспрепятствовать этому. Аменсализмом являются отношения между взрослыми деревьями и всходами деревьев или травами напочвенного покрова в лесу, которые делят ресурсы почвенного питания и азота. В заключение отметим, что взаимоотношения организмов могут меняться не только в зависимости от условий среды (обострение конкуренции при уменьшении ресурсов, кроме того, в разных условиях более сильными конкурентами могут быть разные виды), но и на разных стадиях жизненного цикла. Пример тому – взаимоотношения семги и двустворчатого моллюска жемчужницы. Личинки жемчужницы паразитируют (живут в жабрах семги), но взрослые моллюски живут независимо на дне и, фильтруя воду, улучшают условия для жизни рыбы, очень чувствительной к загрязнению. Кроме того, между скоплениями раковин прячется от хищников молодь семги. Уже описанный положительный «эффект группы» и отношения «растений-нянь» и их подопечных также со временем сменяется острой конкуренцией: в группе начинается процесс самоизреживания, а подопечные подавляют своих «нянь». Положительные взаимодействия играют важную роль в организации экосистем, так как уравновешивают антагонизм конкуренции, хищничества и паразитизма. Контрольные вопросы1. Какие отношения называются комменсализмом? 2. Расскажите об отношениях «растений-нянь» и их «подопечных». 3. Какие взаимоотношения складываются между эпифитами и деревьями. 4. Какую роль играет детритофагия в жизни экосистем? 5. Расскажите о детритофагах, населяющих почву. 6. Какую роль играют копрофаги? 7. Какие организмы называются мусорщиками? 8. Приведите примеры аменсализма. 8.8. Сигнальные взаимоотношения организмов Сигнальные взаимоотношения организмов – это информационные отношения, которые не сопровождаются разделом или передачей от одного организма другому материально-энергетических ресурсов. Они могут достаточно эффективно влиять на результаты горизонтальных (конкуренция) и вертикальных (хищничество, паразитизм) взаимоотношений. Различают несколько типов сигналов. Зрительные сигналы – основа взаимоотношений насекомо-опыляемых растений и насекомых-опылителей, которые опознают растения по яркой окраске венчика. К зрительным сигналам относятся устрашающие позы животных (вздыбленная шерсть, оскал), а также приобретение животными окраски, защищающей от поедания (красный цвет насекомых). К сигналам этого типа относятся и ложные сигналы – мимикрия, когда организмы «подражают» определенным предметам живой или неживой природы. Насекомые могут, к примеру, подражать животным, которые либо несъедобны, либо опасны (безвредный европейский шершневидный мотылек маскируется под жалящего шершня, самки парусника-притворяшки маскируются под некоторых ядовитых бабочек и т.д.). Звуковые сигналы. Такие сигналы подают птицы, оповещающие друг друга о том, что участок занят, или об опасности. Сигналом, предотвращающим стычку хищников, является рычание. Дельфины и некоторые насекомые обмениваются звуковыми сигналами на частотах, недоступных человеческому слуху. Химические сигналы. Этот вид сигнальных отношений особенно широко распространен у животных, которые продуцируют специальные сигнальные вещества – феромоны, обладающие сильным запахом и потому действующие при очень низких концентрациях (половые феромоны, феромоны для мечения территории, агрегационные феромоны – при их посредстве собираются вместе муравьи одной семьи, феромоны тревоги и т.д.). Большую роль играют запахи и во взаимоотношениях насекомых-опылителей и растений. В последние годы описано много фактов влияния химических сигналов растений на интенсивность их поедания фитофагами (см. 6.3.). Более того, выявлены летучие вещества, выделяемые насекомыми во время поедания растения. Эти сигналы играют роль репеллентов, которые отпугивают других насекомых того же вида. Это «выгодно» и растению (снижается скорость его поедания), и насекомому – защищает его от конкуренции. Есть данные, что химические сигналы еще и привлекают энтомофагов и паразитов, которые способны помочь растению справиться с нашествием фитофагов (De Moraes et al., 2001). Сигнальные химические взаимоотношения между растениями называются аллелопатией. Растения выделяют в окружающую среду биологически активные вещества – колины. Они попадают в почву (корневые выделения), в атмосферу (газообразные вещества с сильным запахом) и воду (смывы с листьев). Колины выделяются и при разложении мертвого органического вещества, например лесной подстилки. Считается, что роль колинов во взаимоотношениях растений вторична (т. е. их влияние – это экзаптация), так как первично они вырабатывались растениями для отпугивания насекомых-фитофагов. Факт существования аллелопатии бесспорен и подтвержден в экспериментах сотни раз. Со времен Теофраста известно, что под пологом грецкого ореха не растут другие плодовые культуры. Под интродуцированными на Кавказ австралийскими эвкалиптами из-за аллелопатических выделений из опавших листьев не могут расти местные виды трав. Однако роль аллелопатии в естественных экосистемах невелика, так как в них нет условий, при которых концентрация колинов повышается до уровня, когда они могут существенно влиять на растения. Кроме того, длительно совместно обитающие растения адаптируются к аллелопатическим выделениям друг друга. Возможно, что колины корней играют роль в обеспечении равномерности их распределения в почве, подавая сигнал «занято». Колины, образующиеся при разложении лесной подстилки, ингибируют возобновление многих видов деревьев. Контрольные вопросы1. На какие группы делятся сигнальные взаимоотношения? 2. Расскажите о зрительных сигналах. 3. Приведите примеры звуковых сигналов. 4. Расскажите о разнообразии химических сигналов животнывх. 5. Что такое аллелопатия и какова ее роль в природе? Темы докладов на семинарских занятиях1. Роль и разнообразие конкурентных отношений между видами в природе. 2. Механизмы, смягчающие антагонистические взаимоотношения организмов в экосистеме. 3. Человек как фактор разрушения экологического равновесия взаимоотношений организмов в экосистеме. 4. Возможности человека управлять мутуалистическими отношениями. 5. Соотношение роли материальных и сигнальных взаимоотношений в экосистеме. Глава 9. Экологическая ниша Понятие «экологическая ниша» – краеугольный камень в фундаменте теории современной экологии. И как ни парадоксально, его сложность является причиной отсутствия точного определения. Э. Пианка (1981) пишет о том, что если бы можно было дать точное определение ниши, то экологию можно было бы считать наукой о нишах. О сложности и запутанности проблемы ниши говорит и П. Джиллер (1988). Р. Макинтош (McIntosh, 1974) приводит стихотворение-шутку, которое придумали студенты Корнельского университета. Вот он (в переводе авторов): Когда я слушаю лекции о нише, Стихотворение достаточно точно передает двойственный характер концепции – общетеоретическую значимость для понимания экологии и в то же время ее абстрактный характер, который не дает возможности экологу конкретно определить параметры ниши в природе. 9.1. Экологическая ниша как многомерное явление Экологическая ниша – это совокупность экологических факторов – абиотических и биотических (включая ритмику их изменения) – необходимых для существования популяции в экосистеме. Корни понятия «экологическая ниша» теряются в работах зоологов начала ХХ столетия. Первым его использовал Дж. Гринел (Grinnell, 1914), который понимал под нишей пространство, занимаемое видом в экосистеме (хотя термин «экосистема» в то время еще не был предложен). Спустя 13 лет Ч. Элтон (Elton,1927) использовал понятие «экологическая ниша» для обозначения способа питания вида, т.е. его места в пищевых цепях. Однако наиболее полное представление о нише связано с именем Дж. Хатчинсона (Hutchinson, 1957), который рассматривал как экологическую нишу всю сумму связей вида с абиотическими и биотическими факторами. По Ю. Одуму (1986), экологическая ниша – это «профессия вида в экосистеме» (т.е. из какого сырья он производит органическое вещество, где он работает, каков его рабочий график и кто принимает его продукцию для дальнейшей переработки). Поскольку число факторов, определяющих нишу велико, Хатчинсон предложил «n-мерную модель». В этой модели каждому фактору среды соответствует одна ось абстрактного гиперпространства, т.е. пространства с числом осей больше трех (как в физическом пространстве). И в этом гиперпространстве каждый вид, представленный популяцией, занимает определенный гиперобъем. Гиперобъемы ниш разных видов перекрываются, но не совпадают. Общее число осей гиперпространства ниш определить трудно, и потому в практической деятельности экологи оценивают экологическую нишу по 1-3 осям-факторам, наиболее важным для данного вида. Обычно в качестве таких осей используются ведущие (лимитирующие) экологические факторы (см. 3.3). В литературе описаны десятки примеров разделения (дифференциации) экологических ниш популяциями разных видов по одному из факторов среды. Так, к примеру, два вида морских желудей (Chthamalus stellatus и Balanus balanoides) у побережья Шотландии по разному распределены на литорали: первый вид обитает в ее нижней части при более длительном затоплении, а второй – в верхней при более кратковременном затоплении (Connell, 1961). Ведущей осью разделения экологических ниш является продолжительность затопления. Гиперобъем ниш, разумеется, включает и другие факторы, но для объяснения различий распространения видов оказалось достаточным изучить разделение ниш всего по одной оси – ведущего фактора. Для понимания значения разделения экологических ниш в организации сообществ и экосистем большую роль сыграли эксперименты с инфузориями русского эколога Ф. Гаузе в 30-х гг. ХХ в. Гаузе сформулировал принцип, получивший впоследствии его имя: «Два вида не могут сосуществовать, если они занимают одну экологическую нишу». (Один из них должен погибнуть либо уйти в другую экологическую нишу.) Принцип Гаузе («конкурентное исключение», см. раздел 8.2) в природе действует редко. Поскольку число осей у экологической ниши достаточно велико, то сосуществование видов может быть связано с различиями отношений к одному из второстепенных факторов. Сосуществованию видов способствуют флюктуирующие ресурсы; третий вид, который сдерживает рост конкурирующих популяций; мозаичность местообитания. Во многих случаях одну и ту же нишу поочередно могут занимать несколько видов вследствие «лотереи» (см. 7.2). Нишу захватывает тот вид, который придет первым. После его гибели эту нишу может занять другой претендент, потом третий и т.д. В фитоценологии такую ситуацию описывает «модель карусели» (van der Maarel, Sykes, 1993). Следует отметить, что понятие «экологическая ниша» – достаточно абстрактное. М. Бигон и др. (1989) по этому поводу пишут: «Важно осознать, что экологическая ниша не есть нечто такое, что можно увидеть глазами. Не нужно и вымерять всевозможных ее проекций на отдельные оси – представление о нише сослужит службу и без этого. Экологическая ниша – отвлеченное понятие, сводящее под общий ярлык все, в чем нуждаются организмы, т.е. все те условия среды, которые необходимы им для поддержания жизнеспособной популяции, а также все потребные для этого ресурсы в необходимых количествах. Следовательно, экологическая ниша – это характеристика организма или …вида» (т. 1, с. 110-111). Контрольные вопросы1. Расскажите об истории формирования концепции экологической ниши. 2. Проиллюстрируйте принцип разделения экологических ниш на примерах разных организмов. 3. В каких случаях возможно сосуществование видов, занимающих одну нишу? 9.2. Различия экологических ниш у животных и растений У животных экологические ниши различаются более четко, чем у растений, так как разные животные потребляют разную пищу (биомасса или детрит разных видов растений, насекомых, рыб, птиц или млекопитающих). Животные разделяют территорию на охотничьи наделы или живут в разных стациях одной экосистемы (например разные птицы гнездятся и кормятся в разных частях древесного яруса). Есть животные, которые охотятся днем, а другие – только ночью (филины, совы, летучие мыши, сомы). У перелетных птиц ниши в разные периоды года разные и находятся в разных экосистемах, удаленных друг от друга на тысячи километров. У растений ниши различаются не столь четко, так как у всех растений – одна диета из раствора минеральных солей, который они корнями поглощают из почвы, диоксида углерода и солнечного света. Поэтому до начала 60-х годов понятие экологической ниши применительно к растениям не использовалось, хотя принцип экологической индивидуальности видов (см. 3.2) был известен с конца прошлого столетия. Однако постепенно (в основном благодаря Т.А. Работнову) оно стало обычным и в применении к растениям, так как стало очевидным, что одни и те же ресурсы разные виды растений используют неодинаково. Д. Тильман (Tilman,1988) даже пишет о том, что у каждого растения «свой тип фуражирования». Различия своих «профессий» в экосистеме растения реализуют за счет следующих особенностей (Миркин, Наумова, 1998). Разный тип распределения корневых систем. В пустыне Гоби авторы наблюдали сообщества из тростника и саксаула (Haloxylon ammodendron). Тростник – гигрофит-фреатофит, который связан с грунтовыми водами на глубине 8-10 м, саксаул – типичное пустынное дерево с фотосинтезом САМ и укоренением в сухих горизонтах почвы, из которых он извлекает некоторое количество воды. В высокогорьях Кавказа распространены сообщества пустошей, где лишайники занимают самый верхний слой почвы и используют как источник воды и минеральных элементов атмосферные осадки, а в более глубоком горизонте расположена густая сплошная «сетка» из корней злаков, использующих влагу почвы (Онипченко, 1995). Разное время вегетации. В период цветения и плодоношения растениям требуется больше ресурсов, и за счет цветения в разное время снижается конкуренция. Эта разновременность цветения («экологическая очередь») до неузнаваемости изменяет внешний облик сообществ, что позволяет ботаникам говорить о феномене смены аспектов. Выдающийся исследователь степей В.В. Алехин выделял в степях более 10 аспектов по цветущим в разное время видам. Разная требовательность к свету. В лесу одни виды растений приурочены к затененным местам (например копытень, вороний глаз), а другие – к светлым прогалинам (ежа сборная). Виды подлеска (рябина, лещина, бересклет бородавчатый) довольствуются меньшим количеством света, чем виды высокого древесного яруса. Разная требовательность к элементам минерального питания. Злакам необходим азот, который они получают из почвы. Бобовые могут обходиться практически без него, так как в их корнях живут азотфиксирующие бактерии. Растения различаются также по взаимоотношениям с насекомыми-опылителями, микоризообразующими грибами, фитофагами и другими факторами, которые определяют состав экосистемы. Контрольные вопросы1. В чем состоит основное отличие экологических ниш растений и животных? 2. По каким осям экологических факторов могут дифференцироваться экологические ниши растений. 3. Какие биотические факторы способствуют разделению экологических ниш у растений. 9.3. Фундаментальная и реализованная ниши Разделение экологических ниш у видов одного типа питания никогда не бывает полным, их ниши пересекаются. Например заяц может служить пищей и для лисы, и для волка, но волк, кроме того, может нападать и на более крупных травоядных – оленей и даже лосей, а лиса – использовать в пищу мышевидных грызунов. В фенологической последовательности прохождения фенофаз растениями сравнительно редко цветение одного вида резко сменяется цветением другого, обычно второй вид зацветает в то время, когда первый еще не закончил цветение. Пересекаются распределения видов растений на градиентах ведущих факторов – увлажнения, засоления, освещенности и др. Пересечение экологических ниш является причиной конкуренции, которая, в свою очередь, влияет на размер экологических ниш видов. Это позволило Д. Хатчинсону различать два варианта ниши – фундаментальную и реализованную. Фундаментальная ниша – это ниша, которую вид может занять при отсутствии конкуренции. Она обусловлена генетически. Реализованная ниша – это часть фундаментальной ниши, которую занимает вид при наличии конкуренции. Она обусловлена генетически и экологически. Фундаментальные ниши видов очень широкие. К примеру, в ботанических или зоологических садах при отсутствии конкуренции успешно выращиваются растения и содержатся животные из разных районов земного шара (они защищены от более конкурентоспособных видов местной флоры и фауны). В ботаническом саду университета в Галле в открытом грунте растет опунция американская, но трудно себе представить, чтобы она натурализовалась и внедрилась в травостой злаковников или в сообщества кустарников Южной Германии. В условиях непривычного климата конкурентная способность кактуса крайне низка. Классическими примерами, показывающими различия фундаментальной и реализованной ниш, являются опыты по удобрению естественных лугов и искусственных чистых посевов тех видов, которые слагают эти луговые сообщества (Работнов, 1998). При внесении минеральных удобрений на луга лесной зоны, которые сформировались после расчистки леса на бедных подзолистых почвах, из травостоя выпадают типичные для таких лугов олиготрофные мелкие узколистные злаки – полевица тонкая и душистый колосок и разрастаются эвтрофные крупные широколистные злаки – ежа сборная, коротконожка перистая и пырей ползучий, бывшие ранее второстепенными видами с небольшой долей количественного участия в травостое. Однако и полевица, и душистый колосок в искусственном чистом посеве положительно отзываются на удобрения, внесенные в тех же дозах, и их урожайность увеличивается в 2–4 раза. Причины этого феномена заключаются в том, что олиготрофные виды при отсутствии конкуренции хорошо растут и в условиях богатого минерального питания. Однако в этих же условиях при наличии конкуренции они вытесняются эвтрофными злаками. То есть их фундаментальная ниша охватывает и бедные, и богатые почвы, а реализованная – только бедные, на которых они вследствие биологических и экологических особенностей (патиентности) являются сильными конкурентами. По такой же причине степные злаки – ковыли и типчаки – в чистом посеве при поливе дают урожай выше, чем на сухом степном черноземе. Их реализованная ниша охватывает только условия засушливой степной зоны, где выжить им помогают также черты патиентности – корневая система интенсивного типа (большое количество тонких длинных корней), узкие листья, опушение, С4-фотосинтез и другие приспособления для поглощения и экономного расходования воды. В то же время в отсутствие конкуренции эти степные злаки хорошо растут при обильном увлажнении. Соотношение объемов фундаментальной и реализованной ниш – признак стратегии вида. Так у виолентов они практически равны: эти конкурентно сильные виды занимают любые местообитания, где они могут произрастать в силу своих генетически обусловленных физиологических особенностей. У патиентов объемы реализованных и фундаментальных ниш также совпадают, но уже по иной причине: в экстремальных условиях (сильное засоление, затенение, крайняя бедность или сухость почвы) растительный покров разомкнут и практически нет конкуренции. На скудные ресурсы, которые имеются в этих экстремальных условиях, никто, кроме патиентов (галофитов, сциофитов, олиготрофов или ксерофитов), не претендует. У эксплерентов реализованная ниша приближается к нулю, так как они слабые конкуренты и могут существовать только при условии, что более сильные виды отсутствуют (обычно это и бывает причиной разрастания эксплерентов на местах нарушений). Аналогично соотношение объемов реализованных и фундаментальных ниш у животных, представляющих разные типы стратегии. У видов с вторичными типами стратегий (RS, RC, SC, CRS) соотношение объемов реализованной и фундаментальной ниш может составлять от нуля до единицы в зависимости от того, в каких соотношениях находятся свойства виолентности, патиентности и эксплерентности (чем выше доля эксплерентности, тем оно ближе к нулю). Кроме фундаментальной и реализованной ниш в экологической ботанике используется понятие регенерационной ниши, предложенное П.Граббом (Grubb, 1985). Регенерационная ниша – это набор факторов среды, позволяющий виду внедрится в сообщество. У пойменных тополей и ив регенерационная ниша, к примеру, очень узкая – это свежие и лишенные растительности влажные наносы аллювия в прирусловой пойме. Весьма интересно, что семена этих деревьев оказываются готовыми к прорастанию как раз в то время, когда идет спад уровня паводка и обнажаются влажные супесчаные и суглинистые субстраты. В иных условиях их легкие семена с пушистыми парашютами («тополиный пух») не прорастают, так как зависают между стеблями травянистых растений. Семена, попадающие в густой ивняк, достигают поверхности влажной почвы и прорастают, но всходы гибнут от сильного затенения. Как создание регенерационных ниш можно рассматривать вспашку почвы перед посевом и агротехнический прием поверхностного улучшения – дискования дернины луга перед подсевом в нее ценных видов трав. Контрольные вопросы1. Дайте определения фундаментальной и реализованной ниши. 2. Расскажите об экспериментах, подтверждающих существование реализованных и фундаментальных ниш. 3. Каково соотношение реализованной и фундаментальной ниши у виолентов, патиентов и эксплерентов? 4. Как это соотношение меняется у видов вторичных стратегий? 5. Что такое регенерационная ниша? 9.4. Гильдии К одной гильдии (Джиллер, 1988) относятся виды, которые делят один и тот же ресурс (имеют похожие экологические ниши), и потому потенциально могут быть конкурентами. Ниши видов одной гильдии особенно сильно перекрыты. Примеры гильдий – крупные фитофаги или насекомые-листоеды саванн, мышевидные грызуны в степи, виды растительноядного или плотоядного планктона, детритофаги бентоса, крупные рыбы-хищники и т.д. Как гильдии можно трактовать деревья, образующие лесной полог, или теневыносливые мхи напочвенного покрова. Однако как правило, в использовании термина «гильдия» применительно к растениям необходимости не бывает. Вполне достаточно использовать понятия «жизненная форма» или «экоморфа» (дробная единица классификации жизеннных форм, которая учитывает принадлежность вида к экологической группе, см. 4.5). Гильдия животных – это достаточно удобное понятие для выделения в экосистеме функциональных блоков среди организмов одного трофического уровня. Контрольные вопросы1. Дайте определение гильдии. 2. Приведите примеры гильдий животных. 3. Насколько целесообразно использовать понятие гильдии для растений? Темы докладов на семинарских занятиях1. Значение концепции экологической ниши для современной экологии. 2. Принцип разделения ниш как механизм сосуществования видов в экосистеме. 3. Вклад концепции экологической ниши в развитие представлений о типах стратегий организмов. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Верх |
||||
|
|
||||